Alternative splicing ofregulates plant development under heat stress

专属客服号

微信订阅号
大数据治理
全面提升数据价值
赋能业务提质增效
Abstract
The Phytochrome-Interacting Factor 4 (PIF4) is a key player in the integration of multiple internal and external stimuli to optimize different aspects of plant development. While both the DNA encoding this transcription factor and its protein are known to be under tight control, no regulation at the RNA level has been previously reported. Our genomic analysis revealed that the exon/intron structure of the basic Helix-Loop-Helix (bHLH) DNA binding domain of PIF4 is conserved and pointed to skipping of an exon in this region specifically in response to heat stress. We then showed that this alternative splicing event downregulates PIF4 function under heat, which in etiolated seedlings induces photomorphogenic-related traits. Our results disclose a role for PIFs in plant responses to heat and reveal a new regulatory layer for the control of PIF4 function, underscoring the critical role of posttranscriptional regulatory processes in the molecular integration of environmental cues.
Introduction
Phytochrome Interacting Factors (PIFs) belong to the basic Helix-Loop-Helix (bHLH) family of transcription factors. The bHLH protein domain consists of two segments: the basic region, required for DNA binding, and the helix-loop-helix, responsible for hetero and homodimerization (
Toledo-Ortiz et al., 2003
). PIFs are also characterized by the presence of a protein domain that interacts with photoactivated phytochromes (
Favero, 2020
). This interaction promotes the degradation of PIFs in the light and is crucial for their role as major regulators of light-regulated biological processes (
Leivar and Monte, 2014
). In etiolated seedlings, which germinate and develop in subterranean darkness, PIFs are active and repress photomorphogenic features, such as chlorophyll biosynthesis, cotyledon expansion, and repression of hypocotyl elongation (
Leivar et al., 2008
;
Shin et al., 2009
).
Besides their well-known function in adjusting seedling development to light, PIFs, particularly PIF4, have over the past years also been implicated in the regulation of different biological processes such as immunity (
Gangappa et al., 2017
), morphological adaptations to high ambient temperatures (
Koini et al., 2009
), stomatal development (
Casson et al., 2009
), leaf senescence (
Sakuraba et al., 2014
), freezing tolerance (
Lee and Thomashow, 2012
), salt tolerance (
Wang et al., 2025
), anthocyanin biosynthesis (
Liu et al., 2015
), or fatty acid biosynthesis (
Liao et al., 2025
). PIF4 has thus emerged as a key integrator of multiple external and internal signals to optimize plant development (
Choi and Oh, 2016
;
Lucyshyn and Wigge, 2009
).
Several studies have described the molecular mechanisms that control
PIF4
gene expression, protein levels, and activity (
Leivar and Quail, 2011
;
Paik et al., 2017
;
Pham et al., 2018
), but its posttranscriptional regulation has never been characterized. Here we show that alternative splicing, a posttranscriptional process generating multiple mRNAs from the same gene, produces two different
PIF4
transcripts specifically in response to heat stress.
Temperature deviations from the optimal range significantly impact plant development and survival. Increases in temperature are classified as either high ambient temperature or excessively hot temperatures. High ambient temperature is typically 5-6ºC above the optimum temperature (22ºC for
Arabidopsis thaliana
), while excessively hot temperatures exceed this range (
Li et al., 2018
). These distinct temperature ranges activate independent signaling pathways, leading to different physiological outcomes. Warm temperatures induce thermomorphogenesis, which generally promotes growth and development in a PIF4-dependent manner (
Quint et al., 2016
). Conversely, excessively hot temperatures trigger stress-responsive pathways aimed at adjusting growth and physiology to mitigate the negative effects of heat (
Kan et al., 2023
). To date, the role of PIFs in temperature signaling has centered on thermomorphogenesis, with only a few studies having explored the role of PIF proteins in heat stress responses (
Li et al., 2021
;
Yang et al., 2022
). Intriguingly, our results reveal that heat stress induces photomorphogenic features in etiolated seedlings and that this developmental response is mediated by an alternative splice form of
PIF4
.
Results
PIF4
is alternatively spliced in response to heat stress
Our analysis of the exon and intron positioning in all 15 members of the XV subfamily of
Arabidopsis thaliana
bHLH transcription factors that includes PIFs (
Toledo-Ortiz et al., 2003
) showed that, with the sole exception of the less conserved member HFR1, the intron/exon distribution of the bHLH domain is maintained. Its 150-bp long sequence is distributed among three exons, following an invariable proportion: 22%, 44% and 34% (
Figure 1A
). In addition, the middle exon containing the largest section of the bHLH domain is always 66-bp long and surrounded by phase 0 introns, which preserve codon identities and therefore the reading frame (
Figure 1A
). Hence, alternative splicing of this exon will produce protein isoforms differing only in the bHLH region (Supplemental Figure 1). Given the genomic particularities of the bHLH middle exon in these genes, we investigated its splicing regulation by quantifying its PSI (Percent Spliced In; percentage of transcripts that include the exon) in publicly available
Arabidopsis thaliana
RNA-seq samples covering several environmental conditions and tissues at different developmental stages (Supplemental Table 1). This analysis revealed skipping of the exon exclusively in two genes:
PIF4
and
PIF6
(
Figure 1B
and Supplemental Table 1). In the case of
PIF6
, although exon skipping occurs in nearly all tissues and conditions tested, this alternative splicing event has only been shown to be functional in seeds and embryos (
Penfield et al., 2010
), where
PIF6
is highly expressed (Supplemental Figure 2). Strikingly, we found that the
PIF4
gene undergoes alternative splicing in this particular exon, exon 5, exclusively when plants are under heat stress (
Figure 1B
and Supplemental Figure 3). Given that the protein arising from this exon skipping event will lack a portion of the bHLH domain (Supplemental Figure 1), we expect heat-induced alternative splicing to reduce the amounts of functional PIF4.

Alternative splicing regulation of the bHLH central exon in the XV subfamily of Arabidopsis
thaliana
bHLH transcription factors.
(A)
Exon and intron gene distribution (left) and phylogeny (right; adapted from
Leivar and Quail, 2011
) of all members of the XV subfamily of Arabidopsis bHLH transcription factors. The bHLH subdomains are shown in blue. For all genes with the exception of
HFR1
, percentages indicate the proportion of the bHLH domain encoded in each exon.
(B)
PSI (Percent Spliced In) of the bHLH central exon using publicly available RNA-seq data covering several
Arabidopsis thaliana
tissues and environmental conditions (Supplemental Table 1). Blue dots indicate PSI<90, the generally considered cutoff for exon skipping. n.d., not detected.
Heat stress induces photomorphogenesis in the dark
To study the impact of the exon skipping event in the bHLH domain of
PIF4
, we applied heat stress (37 ºC) to 3-day-old etiolated seedlings, a developmental stage at which PIFs are known to be functional in repressing photomorphogenesis (
Leivar et al., 2008
;
Shin et al., 2009
). First, we confirmed that the heat-induced
PIF4
alternative splicing event occurs in etiolated seedlings as well (
Figure 2A
), also showing that it does not occur in response to light and is sustained along time (
Figure 2A
and
2B
). Remarkably, heat treatment of dark-grown seedlings partially induces photomorphogenesis — cotyledons open and hypocotyl elongation is repressed (
Figure 2C
and
2D
). These morphological changes, although less pronounced, are characteristic of seedlings lacking PIF activity, as is the case with etiolated seedlings transferred to light or dark-grown quadruple
pif1pif3pif4pif5
(
pifq
) mutants (
Figure 2B
and Supplemental Figure 4) (
Leivar et al., 2009
,
Leivar et al., 2008
;
Shin et al., 2009
). Moreover, we quantified protochlorophyllide (Pchlide), the phototoxic chlorophyll precursor that, when overaccumulated in etiolated seedings, leads to photobleaching upon light exposure. This analysis revealed higher levels of Pchlide and increased photobleaching in wild-type (WT) etiolated seedlings exposed to heat for 24 hours prior to light exposure, a trend also partially phenocopying
pifq
mutants (
Figure 2E, 2F
and Supplemental Figure 5) (
Leivar et al., 2009
). These phenotypes are consistent with heat-induced alternative splicing reducing the amounts of functional PIF4. Analysis of
PIF1, PIF3, PIF4
and
PIF5
expression levels under 37ºC demonstrated that none of these genes were transcriptionally downregulated by heat stress (
Figure 2G
and Supplemental Figure 6), discarding a strong reduction in
PIF
transcript levels as the cause of the observed phenotypes. Heat-stressed etiolated seedlings are phenotypically more similar to higher order
pif
mutants than to
pif4
single mutants (Supplemental Figure 7 and 8) (
Leivar et al., 2008
;
Shin et al., 2009
), suggesting that the
PIF4-S
splice form generated by exon skipping may act as a dominant negative rather than being merely inactive. Previous studies have reported that different PIF4 protein isoforms can exert dominant negative effects, inhibiting the activity of other PIFs, and that alternative splicing can produce dominant negative transcription factor isoforms (
Gangappa et al., 2017
;
Kim et al., 2020
;
Nicolas et al., 2015
;
Seo et al., 2011
). However, direct evidence is needed to confirm that the
PIF4
short isoform functions as a dominant negative, reducing the activity of the long isoform and other PIFs. Nevertheless, because heat-induced phenotypes are milder than those observed in
pifq
seedlings (
Figure 2C, 2E
and
2F
), a substantial fraction of PIFs likely remains functional at 37 ºC. This is consistent with heat-induced alternative splicing affecting only around 50% of transcribed
PIF4
mRNAs (
Figure 2A
and
2B
).
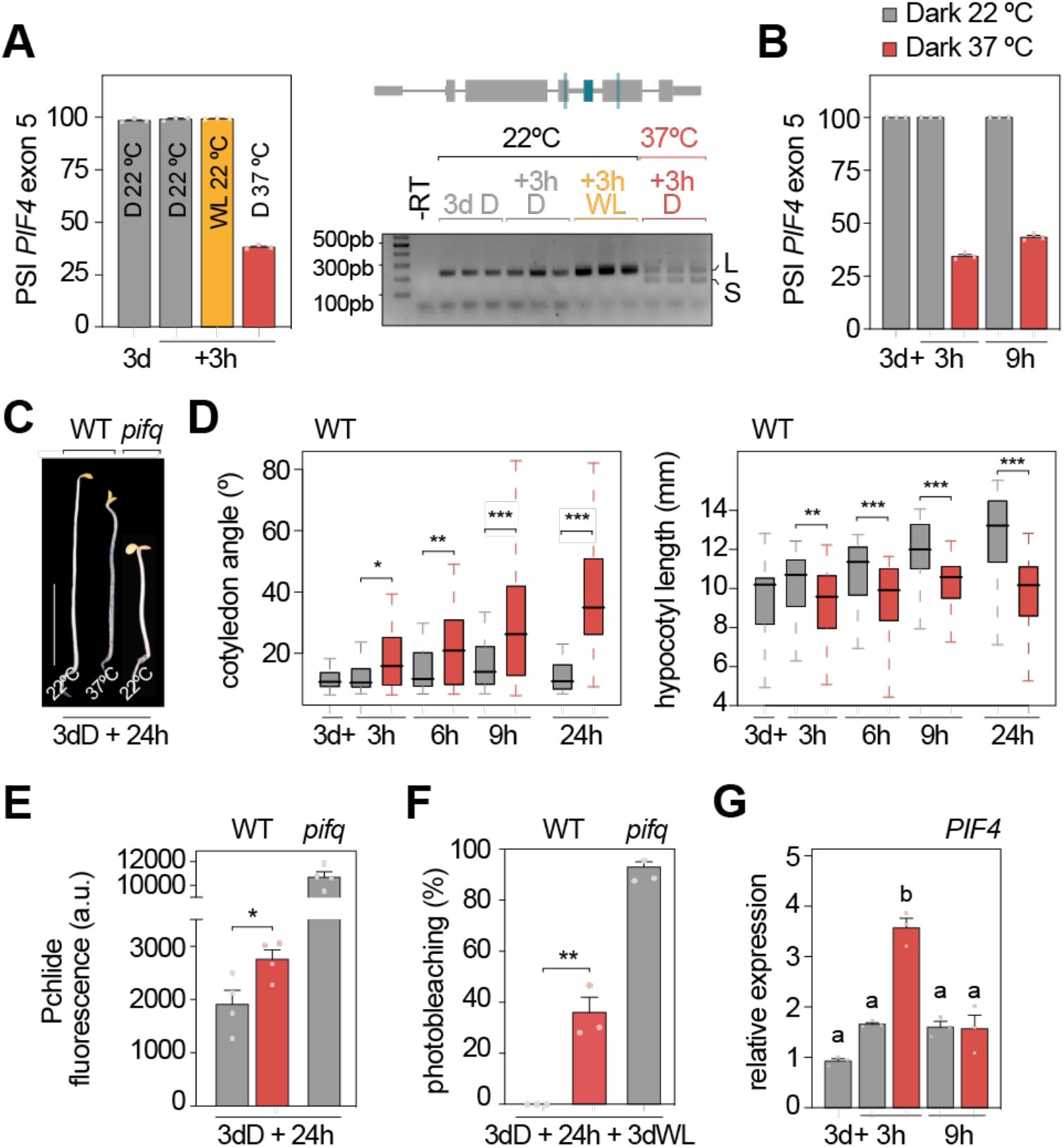
Impact of heat treatment in etiolated seedlings.
(A-B)
PSI (Percent Spliced In) quantification (left) from RT-PCR (right) of the
PIF4
alternatively-spliced exon in seedlings grown in continuous dark for 3 days (d) and then transferred to 37 ºC (red) or white light (WL; yellow), or kept at 22 ºC for 3 (A) or 9 (B) additional hours (h) in darkness (D; gray). Data represent means ± SEM of biological triplicates. -RT, no reverse transcriptase.
(C)
Representative image of 3-day-old wild-type (WT) and
pifq
seedlings subjected or not to heat stress. Scale bar, 5 mm.
(D)
Boxplot representations of the cotyledon aperture (left) and hypocotyl length (right) of at least 35 WT seedlings grown in darkness for 3 days and then transferred to 37 ºC in darkness (red) or kept in the dark at 22 ºC (gray) for the indicated time in hours. Asterisks indicate statistically differences between medians (Mann Whitney test). Fluorescence of protochlorophyllide (Pchlide; 635 nm)
(E
;
n
=4) and percentage of photobleaching
(F
;
n
=3) in heat-stressed (red) or unstressed (gray) etiolated WT and
pifq
seedlings. Asterisks indicate statistically differences between averages (
t
-test).
(G)
RT-qPCR analysis of
PIF4
transcript levels in WT seedlings grown as in (
B
). Values were normalized to
PP2A
and expression levels are expressed relative to the initial time point set at one. Data represent means ± SEM of biological triplicates, and different letters denote statistically significant differences under each condition (Tukey test;
P
<0.05). n.d., not detected. (A, D, E, F) *
P
<0.05, **
P
<0.01 and ***
P
<0.001.
Heat-induced photomorphogenesis depends on
PIF4
alternative splicing
To confirm the implication of
PIF4
alternative splicing in the physiological changes undergone by heat-stressed etiolated seedlings, we quantified the morphological and chlorophyll-related phenotypes of transgenic plants expressing predominantly the long
PIF4
splice form in the
pif4-101
mutant background, (
PIF4p::PIF4-L
;
Figure 3A
and Supplemental Figure 9). Importantly, both the heat-induced Pchlide accumulation and cotyledon opening were strongly reduced in these lines, while the repression of hypocotyl elongation was maintained (
Figure 3B, 3C
and Supplemental Figure 10). In agreement with the reduction of Pchlide in
PIF4-L
plants, a non-significant but correlating trend in their photobleaching phenotypes was also observed (
Figure 3B
). Importantly,
PIF4-L
.
1
expresses the long isoform at levels similar to those of WT plants (Supplemental Figure 9A), ruling out the possibility that the suppression of heat-induced phenotypes (cotyledon opening and Pchlide accumulation) is due to elevated
PIF4
expression levels. In addition, consistent with the comparable alternative splicing levels observed in heat-stressed WT and
pif4
seedlings (
Figure 3A
and Supplemental Figure 9B), the skipped exon is located upstream of the
pif4-101
mutation (Supplemental Figure 9C), and the phenotypes are also comparable (
Figure 3
). Similar results were obtained with the other commonly used
pif4
mutant (
pif4-2
;
Leivar et al., 2008
), which harbors a similar insertion site (Supplemental Figure 9D). Overall, our results substantiate a role for
PIF4
alternative splicing in controlling heat-induced developmental responses in etiolated seedlings. Next, we quantified the transcriptional response to heat stress in etiolated seedlings with different levels of the long
PIF4
splice form (
Figure 3A
). Interestingly, we found a strong enrichment of heat-regulated genes among those reported as PIFq-regulated or PIFq-bound (
Pfeiffer et al., 2014
) (
Figure 3D
). Furthermore, heat-induced transcriptional changes in
pif4
mutants, the genetic background of
PIF4-L
seedlings, were significantly attenuated in these transgenic lines, yet the response remained far from abolished (
Figure 3E
). This result could be explained by some heat-induced transcriptional changes being fully PIF-independent, as shown in
Figure 3D
, and others being PIF-dependent but unaffected due to the considerable fraction of PIFs still functional under heat stress. Either scenario would also explain the partial reversion of the heat-induced phenotypic responses observed in
PIF4-L
lines.
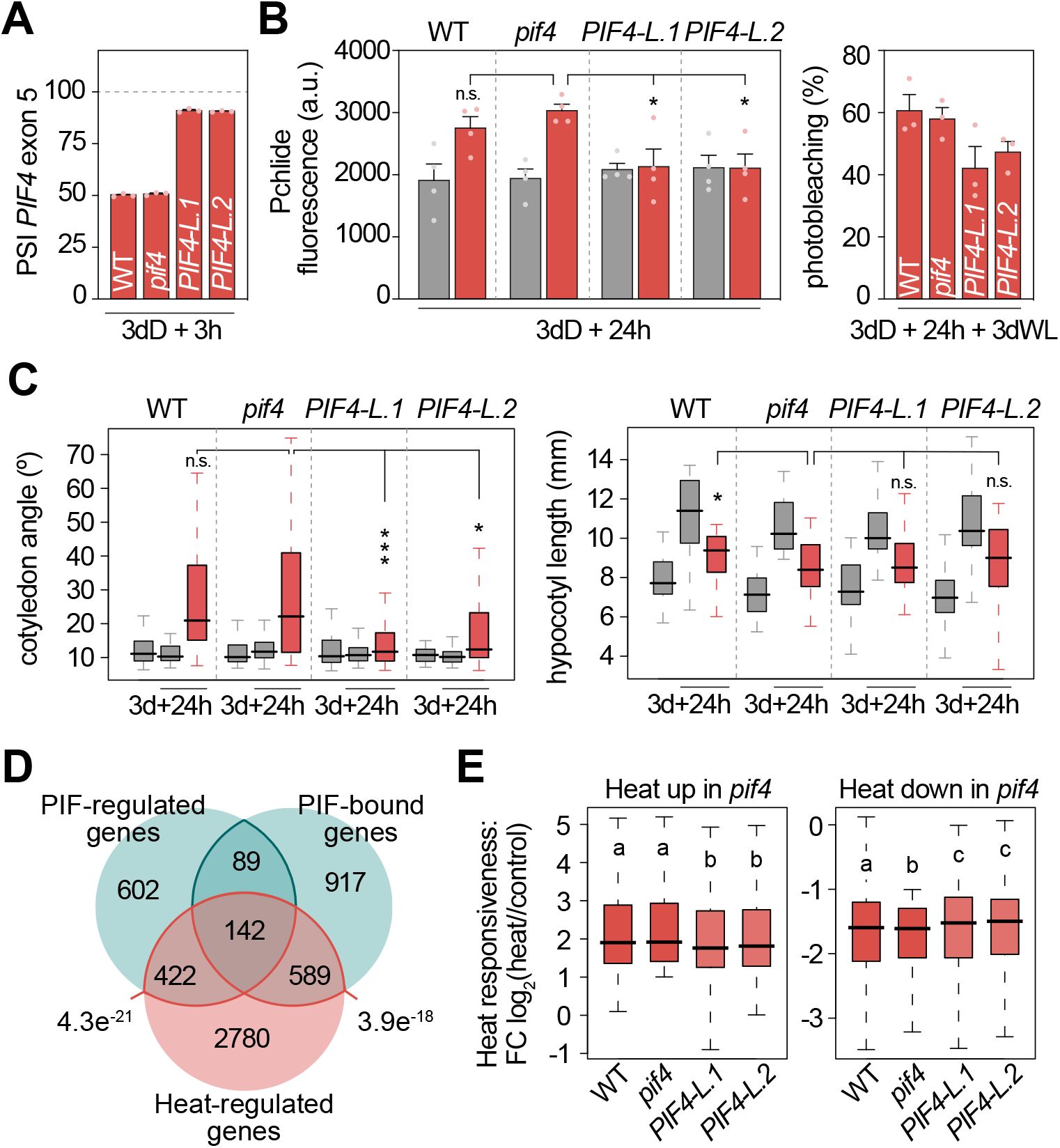
Enhancing expression of the
PIF4
long isoform at 37ºC reduces the impact of heat stress in etiolated seedlings.
(A)
PSI (Percent Spliced In) quantification of the
PIF4
alternatively-spliced exon in wild-type (WT),
pif4
and
PIF4-L
seedlings grown in continuous dark for 3 days (d) and then transferred to 37 ºC for 3 additional hours (h) in darkness. Data represent means ± SEM of biological triplicates.
(B)
Protochlorophyllide (Pchlide; 635 nm) fluorescence (left;
n
=4) and percentage of photobleaching (right;
n
=3) in 3-day-old etiolated WT,
pif4
and
PIF4-L
seedlings transferred to 37 ºC (red) or kept at 22 ºC (gray) for 24 additional hours (h) in the dark. For photobleaching quantification, seedlings were subsequently exposed to white light (WL) for 3 days. Data represent means ± SEM of biological replicates (t-test). a.u., arbitrary units.
(C)
Boxplot representations of the cotyledon aperture (left) and hypocotyl length (right) of at least 35 WT,
pif4
and
PIF4-L
seedlings grown as in (
B
). Mann Whitney test was used to define statistically differences.
(D)
Venn diagram showing overlap among heat-regulated genes in WT seedlings defined in this study and PIF-regulated and PIF-bound genes defined previously (
Pfeiffer et al., 2014
) (two-sided Fisher’s exact test).
(E)
Heat responsiveness (fold change; FC) in WT,
pif4
and
PIF4-L
for heat-regulated genes in
pif4
seedlings (
n
=3). Different letters denote statistically significant differences between genotypes by Dunn’s test (
P
<0.05). (A-C) Asterisks indicate statistically significant differences from
pif4
(*
P
<0.05, **
P
<0.01 and ***
P
<0.001; n.s., not significant), and
n
the number of biological replicates.
To confirm the role of
PIF4
alternative splicing in regulating heat-induced responses, we generated transgenic plants expressing the short isoform of
PIF4
under the control of its endogenous promoter (
PIF4p::PIF4-S
) and evaluated their morphology in the dark under control temperature conditions. These transgenic lines (
PIF4-S
) showed higher
PIF4
expression levels than the corresponding WT control (
Figure 4A
), and in all three lines, the short isoform was the predominantly expressed variant (
Figure 4B
). We then conducted a phenotypic analysis of seedlings grown in the dark for 3 and 4 days at 21ºC. Interestingly, our results showed that enhanced production of the short isoform consistently promoted cotyledon opening, while changes in the hypocotyl length were not always detectable (
Figure 4C
). Thus, the cotyledon phenotype of these plants resembles that of WT plants exposed to heat stress (
Figure 2C
and
2D
), linking the production of this isoform with heat-induced morphological adaptations. Notably, cotyledon opening in these transgenic plants at 21ºC is less pronounced than in heat-stressed plants (37ºC), indicating that the production of this isoform is not the unique mechanism underlying heat-induced cotyledon opening.
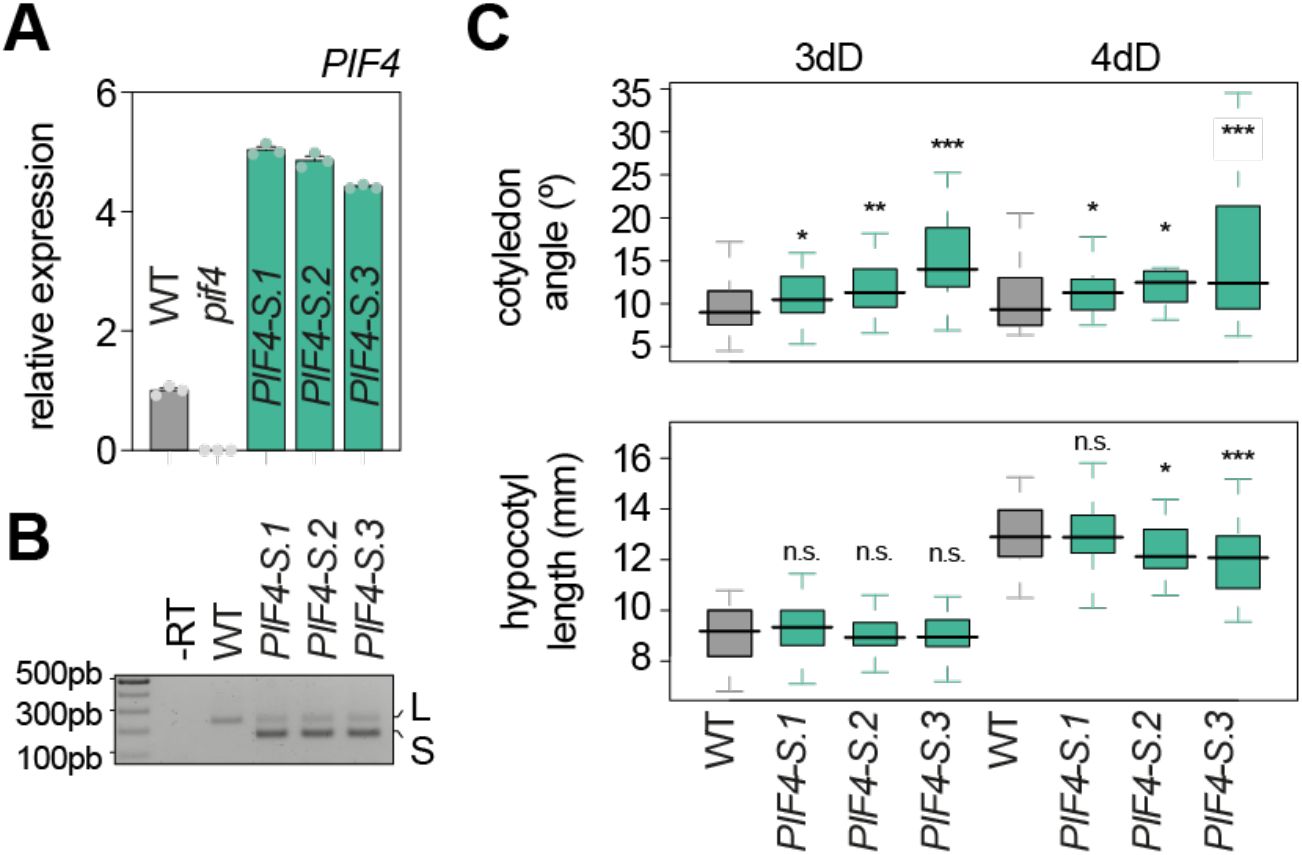
Enhancing expression of the
PIF4
short isoform promotes cotyledon opening in the dark.
(A)
RT-qPCR analysis of
PIF4
transcript levels in wild-type (WT),
pif4
and
PIF4-S
seedlings grown for 4 days in darkness. Values were normalized to
PP2A
and expression levels are expressed relative to the WT. Data represent means ± SEM of technical triplicates.
(B)
RT-PCR of the alternatively-spliced exon of
PIF4
in seedlings grown as in (
A
). -RT, no reverse transcriptase.
(C)
Boxplot representations of the cotyledon aperture (top) and hypocotyl length (bottom) of at least 28 WT,
pif4
and
PIF4-S
seedlings grown for 3 or 4 days (d) in the dark (D). Asterisks indicate statistically differences from WT at each day (Mann Whitney test; *
P
<0.05, **
P
<0.01 and ***
P
<0.001; n.s., not significant).
Discussion
Our study reveals, for the first time, that cotyledon opening is a developmental response of etiolated seedlings exposed to heat stress. Heat stress also exerts a repressive effect on hypocotyl elongation in etiolated seedlings, a phenomenon previously reported but not extensively studied (
Hong and Vierling, 2000
;
Karayekov et al., 2013
;
Larkindale et al., 2005
;
Martín and Duque, 2022
). Karayekov et al. linked this inhibitory effect on hypocotyls to altered functioning of light signaling components such as CONSTITUTIVE PHOTOMORPHOGENIC 1 (COP1), ELONGATED HYPOCOTYL5 (HY5) and circadian clock components. Notably, they also proposed a physiological rationale for this response: dark-germinated seedlings approaching the soil surface may become more susceptible to heat shock episodes due to their proximity to sunlight. They hypothesized that the molecular mechanisms triggered by these episodes would prime seedlings for imminent light exposure. Given that the first exposure to sunlight represents a critical phase for etiolated seedlings, requiring rapid adaptation to ensure survival, we agree that the early activation of photomorphogenesis-associated traits could be advantageous. Our finding that heat promotes cotyledon opening, another hallmark of photomorphogenesis (
Arsovski et al., 2012
), supports the hypothesis that, in etiolated seedlings, heat may function as a signal to induce photomorphogenesis. In addition, we show that heat enhances Pchlide accumulation in dark-grown seedlings. Although the conversion of Pchlide to chlorophyllide is light-dependent, Arabidopsis seedlings accumulate Pchlide in the dark to expedite this process upon light exposure (
Sperling et al., 1997
). However, the amount of Pchlide must be tightly balanced with the availability of the enzyme that catalyzes its conversion to prevent the generation of reactive oxygen species and subsequent cellular damage upon illumination (
Mochizuki et al., 2010
;
Reinbothe et al., 1996
). This raises the question of whether heat-induced Pchlide accumulation in etiolated seedlings is an adaptative mechanism to accelerate chlorophyll production and optimize the transition to autotrophic development, or whether it is a side effect of prematurely activating light signaling in the dark, an outcome that, as suggested by increased photobleaching, would negatively impact seedling survival.
Our data also indicate that the induction of these photomorphogenic traits in etiolated seedlings depends, at least in part, on a heat-specific regulatory event: the alternative splicing of
PIF4
. This represents a novel finding, as PIF4, despite being one of the most studied proteins in Arabidopsis, has been reported to be regulated only at the transcriptional and post-translational levels (
Balcerowicz, 2020
;
Favero, 2020
). Notably, our phenotypic analyses of seedlings with altered patterns of
PIF4
alternative splicing (
PIF4-L
and
PIF4-S
) suggest that the heat-induced isoform plays a role specifically in controlling cotyledon-related phenotypes. This implies that the mechanisms reported by Karayekov to control hypocotyl elongation under heat stress may operate in parallel with the alternative isoform of
PIF4
. Because our transcriptomic experiment was conducted using whole seedlings, we were unable to assess organ-specific effects in detail. We hypothesize that this alternative splicing event may be organ-specific or, alternatively, that the protein encoded by the heat-induced
PIF4
isoform may be preferentially active in cotyledons due to specific protein interactors and/or molecular targets. Further research into organ-specific dynamics is needed to elucidate why this alternative splicing event appears to predominantly affect cotyledon development. These findings would provide valuable insights into the organ-specific roles of PIF proteins, an emerging area of research (
Dong et al., 2019
;
Sun et al., 2016
;
Zhang et al., 2021
).
Our study focuses on the role of PIF4 under heat stress, a condition in which its function remains poorly understood. In fact, the few previous reports linking PIF function to heat stress have yielded contrasting results, with PIFs acting as either positive (
Li et al., 2021
) or negative (
Yang et al., 2022
) regulators of the response. Here, we significantly expand the current understanding of this transcription factor by demonstrating its involvement in heat stress and and further reinforcing its role as a key integrator of diverse environmental signals in the regulation of plant development (
Paik et al., 2017
). Importantly, modulating alternative splicing to alter isoform abundance is emerging as a promising strategy for developing stress-resilient plants (
Alhabsi et al., 2025
). In this context, investigating the role of the
PIF4
alternative isoform in heat-stressed adult plants, along with molecular strategies that specifically target the splicing sites involved in its regulation, could reveal a novel molecular target and offer an alternative genetic approach to enhance plant stress tolerance.
Methods
Plant materials
The
Arabidopsis thaliana pif1-1pif3-3pif4-2pif5-3
(
pifq
) and
pif4-2
mutant was obtained from the Nottingham Arabidopsis Stock Centre (NASC).
PIF4-L
transgenic plants expressing the
PIF4
coding region driven by the endogenous promoter, together with its respective
pif4
mutant control (
pif4-101
;
Lorrain et al., 2008
) were kindly provided by U. Pedmale (Cold Spring Harbor Laboratory, USA).
PIF4-S
transgenic plants were generated by PCR amplification and cloning a 1227-bp fragment containing the coding sequence region of the short splice variant under the control of a 2505-bp fragment upstream the PIF4 start codon corresponding to the PIF4 promoter, in the eGFP-tagged version of the binary pBA002 vector using the XbaI/AatI restriction sites (5’-GACGTT
TCTAGA
ATGGAACACCAAGGTTGGAG
-3’and 5’-GT
GACGTC
CGAGTGGTCCAAACGAGAAC
-3’). The pPIF4 promoter was insertved into the promoterless pBA002 via HindIII/XbaI restriction sites (5’-
TGTGAAGCTTCCAAAGTAATAAAAGTTGCCACAAC
-3’and 5’-
GACGTTTCTAGAGTCAGATCTCTGGAGACATTTC
-3’). The resulting constructs were introduced into
Agrobacterium tumefaciens
strain EHA105 and subsequently used for agroinfiltration-mediated transformation of Col-0 seedlings (
Clough and Bent, 1998
).
Phenotypical and photobleaching analysis
Sterile seeds were sown on MS medium containing 1X Murashige and Skoog (MS) salts (Duchefa Biochemie), 2.5 mM MES (pH 5.7), 0.5 mM myo-inositol, and 0.8% agar (w/v). After stratification for 4 days at 4 ºC in darkness, seeds were subjected to a 3-hour light pulse to induce germination and then transferred to continuous darkness for 69 hours at 22 ºC. Maintaining the absence of light, seedlings were then either kept at 22 ºC for control conditions or transferred to 37 ºC for heat stress. Hypocotyl and cotyledon measurements of at least 30 seedlings and two biological replicates were carried out using the National Institutes of Health ImageJ software as described before (
Sentandreu et al., 2011
). Pictures were taken before and after exposure to stress as indicated in each figure. Photobleaching experiments were adapted from previous studies (
Leivar et al., 2009
), with 3-day-old etiolated seedlings being grown under control conditions or at 37 ºC for 24 hours and then transferred to continuous white light for 3 additional days (100 μmol·m
−2
·s
−1
). At this point, the percentage of seedlings that failed to become green were scored in three biological replicates.
Protochlorophyllide quantification
Approximately 30 sterile seeds, sown on MS medium and stratified for 4 days at 4 ºC in the dark, were subjected to a 3-hour light pulse before being transferred to continuous darkness for 69 hours at 22 ºC. Seedlings were then either kept under these conditions (control) or transferred to 37 ºC in the dark to induce heat stress. Whole seedlings were collected in the dark 24 hours later, flash-frozen in liquid N
2
,and ground before extraction with 0.75 mL ice-cold 9:1 acetone:0.1 M NH4OH, as described previously (
Terry and Kacprzak, 2019
). The resulting mixture was vortexed for 1 minute and then centrifuged at 14,000 rpm at 4 ºC for 5 minutes. After supernatant recovery, the protochlorophyllide (Pchlide) content was determined as the peak value (635 nm) of the fluorescence emission spectrum between 600-700 nm, measured with a bandwidth of 5 nm after excitation at 440 nm and using a Synergy Neo2 microplate reader (Biotek). Pchlide data is shown as the average of Pchlide per seedling of four biological replicates.
Gene expression and PSI quantification from RNA extraction
Total RNA was extracted from
Arabidopsis thaliana
seedlings using the InnuPREP Plant RNA kit (Analytik Jena BioSolutions) and 1 µg treated with DNase I to remove genomic DNA. cDNA synthesis using the oligo dT primer and the enzyme SuperScript III reverse transcriptase (Invitrogen) was conducted in the presence of RNase Out (Invitrogen). The cDNA was then used to quantify either gene expression or exon skipping of
PIF4
’s fifth exon. In both cases, three biological replicates were analyzed for each condition and/or genotype tested. Gene expression was measured by Reverse Transcription-quantitative PCR (RT-qPCR) using a QuantStudioTM 7 Flex Real-Time PCR System 384-well format and the Absolute SYBR Green ROX Mix (Thermo Scientific) on 2.5 µL of cDNA (diluted 1:10) per 10 µL of reaction volume, containing 300 nM of each gene-specific primer (see below). The
PP2A
gene was used for normalization (
Shin et al., 2007
). Exon skipping of the fifth exon of
PIF4
was quantified from RT-PCRs using primers spanning the two adjacent exons. These primer sequences were obtained from PastDB (Plant alternative splicing and transcription Data Base;
www.pastdb.crg.eu
;
Martín et al., 2021
). The resulting bands were quantified using the National Institutes of Health ImageJ software.
RNA sequencing
RNA was extracted from 3-day-old WT,
pif4-101, PIF4-L
.
1
and
PIF4-L
.
2
etiolated seedlings grown for 3 hours at 37 ºC or 22 ºC in complete darkness. Oligo dT, non-strand specific libraries from triplicate biological replicates were built and sequenced using NextSeq500 at the Gulbenkian Institute for Molecular Medicine (GIMM). An average of 15 million 75-nucleotide single-end reads were generated per sample. Raw sequencing data was submitted to the Sequence Read Archive (accession number GSE200247).
Gene expression quantification from RNA sequencing data
Quantification of
Arabidopsis thaliana
transcript expression from our RNA-seq experiment (GSE200247) and public sequencing data (Dataset S1) was performed using vast-tools v2.5.1 and v2.2.2 (
Martín et al., 2021
;
Tapial et al., 2017
), respectively. For each Arabidopsis transcript, this tool provides the number of mapped reads per million mapped reads divided by the number of uniquely mappable positions of the transcript (cRPKM; corrected-for-mappability reads per kbp of mappable sequence per million mapped reads) (
Labbé et al., 2012
). To identify genes differentially expressed between different temperatures, we used vast-tools compare_expr using the option -norm to perform a quantile normalization of cRPKM values between samples. Next, we filtered out the genes that were not expressed at cRPKM > 5 and had read counts > 50 across all the replicates of at least one of the samples compared. Finally, differentially-expressed genes were defined as those with a fold change of at least 2 between each of the individual replicates from each genotype. See
https://github.com/vastgroup/vast-tools
for details.
PSI quantification from RNA sequencing data
We employed vast-tools v2.2.2 to quantify alternative splicing from public sequencing data (
Martín et al., 2021
;
Tapial et al., 2017
). This tool quantifies exon skipping (ES), intron retention (IR) and alternative donor (ALTD) and acceptor (ALTA) site choices. For all these types of events, vast-tools estimates the Percent Spliced In (PSI) of the alternative sequence using only exon-exon (or exon-intron for IR) junction reads and provides information about the read coverage See
https://github.com/vastgroup/vast-tools
for details. Data shown in
Figure 1
and Supplemental Table 1 indicate the PSI quantification of specific alternative splicing events in the subfamily XV of the bHLH transcription factors (see below) using a wide array of samples (Supplemental Table 1).
Data availability
RNA-seq data have been deposited in Gene Expression Omnibus (GEO) (GSE200247).
Acknowledgements
We thank U. Pedmale for kindly providing
pif4-101
mutants and
PIF4pro:PIF4-3xFlag
transgenic lines, and V. Nunes for excellent plant care at the Gulbenkian Institute for Molecular Medicine (GIMM) Plant Facility. This work was funded by Fundação para a Ciência e a Tecnologia (FCT) through grants PTDC/BIA-FBT/31018/2017, PTDC/BIA-BID/30608/2017 and PTDC/ASP-PLA/2550/2021 as well as by the Spanish Ministry of Science and Innovation trough grant PID2021-125223NA-I00 (MCIN/AEI/10.13039/501100011033/FEDER). Funding from the research unit GREEN-it “Bioresources for Sustainability” (ID/04551/2025, UID/PRR/04551/2025) and the Generalitat de Catalunya (AGAUR, GRE2021, ref. SGR00873) is also acknowledged. G.M. was supported by an EMBO Long-Term Fellowship (ALTF 1576-2016), a Marie Skłodowska-Curie Individual Postdoctoral Fellowship (EU project 750469) and a Ramón y Cajal fellow from the Spanish Ministry of Science and Innovation (RYC2021-032539-I). T.L was supported by a Marie Skłodowska-Curie Individual Postdoctoral Fellowship (EU project 706274).
Additional information
Author contributions
M.N.-G., B.A., D.S., T.L. and G.M performed the experiments and analyzed the data. All authors discussed the results. G.M. conceived the project and designed research. G.M. and P.D. wrote the manuscript.
Funding
FCT - Fundação para a Ciência e a Tecnologia (PTDC/BIA-BID/30608/2017)
Guiomar Martín
FCT - Fundação para a Ciência e a Tecnologia (PTDC/BIA-FBT/31018/2017)
Paula Duque
FCT - Fundação para a Ciência e a Tecnologia (PTDC/ASP-PLA/2550/2021)
Paula Duque
FCT - Fundação para a Ciência e a Tecnologia (ID/04551/2025,UID/PRR/04551/2025)
Paula Duque
Spanish Ministry of Science and Innovation (PID2021-125223NA-I00)
Guiomar Martín
EMBO (ALTF 1576-2016)
Guiomar Martín
Spanish Ministry of Science and Innovation (RYC2021-032539-I)
Guiomar Martín
MSCA-IF European Commission (750469)
Guiomar Martín
MSCA-IF European Commission (706274)
Tom Laloum
Additional files
Supplementary file.
Supplemental Figures 1-10 and Supplemental Tables 1-2.
References
Alternative Splicing Dynamics in Plant Adaptive Responses to Stress
Annual Review of Plant Biology
76
:687–717
https://doi.org/10.1146/annurev-arplant-083123-090055
Google Scholar
Photomorphogenesis
The Arabidopsis Book
10
:e0147–e0147
https://doi.org/10.1199/tab.0147
Google Scholar
PHYTOCHROME-INTERACTING FACTORS at the interface of light and temperature signalling
Physiologia Plantarum
169
:347–356
https://doi.org/10.1111/ppl.13092
Google Scholar
phytochrome B and PIF4 Regulate Stomatal Development in Response to Light Quantity
Current Biology
19
:229–234
https://doi.org/10.1016/j.cub.2008.12.046
Google Scholar
PIF4 Integrates Multiple Environmental and Hormonal Signals for Plant Growth Regulation in <italic>Arabidopsis</italic>
Molecules and Cells
39
:587–593
https://doi.org/10.14348/molcells.2016.0126
Google Scholar
Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana
The Plant Journal
16
:735–743
https://doi.org/10.1046/j.1365-313x.1998.00343.x
Google Scholar
The Transcription Factors TCP4 and PIF3 Antagonistically Regulate Organ-Specific Light Induction of SAUR Genes to Modulate Cotyledon Opening during De-Etiolation in Arabidopsis
The Plant cell
31
:1155–1170
https://doi.org/10.1105/tpc.18.00803
Google Scholar
Mechanisms regulating PIF transcription factor activity at the protein level
Physiologia Plantarum
169
:325–335
https://doi.org/10.1111/ppl.13075
Google Scholar
PIF4 Coordinates Thermosensory Growth and Immunity in Arabidopsis
Current Biology
27
:243–249
https://doi.org/10.1016/j.cub.2016.11.012
Google Scholar
Mutants of Arabidopsis thaliana defective in the acquisition of tolerance to high temperature stress
Proceedings of the National Academy of Sciences
97
:4392–4397
https://doi.org/10.1073/pnas.97.8.4392
Google Scholar
The molecular basis of heat stress responses in plants
Molecular Plant
16
:1612–1634
https://doi.org/10.1016/j.molp.2023.09.013
Google Scholar
Heat Shock– Induced Fluctuations in Clock and Light Signaling Enhance Phytochrome B– Mediated Arabidopsis Deetiolation
The Plant Cell
25
:2892–2906
https://doi.org/10.1105/tpc.113.114306
Google Scholar
The epidermis coordinates thermoresponsive growth through the phyB-PIF4-auxin pathway
Nature Communications
11
:1053
https://doi.org/10.1038/s41467-020-14905-w
Google Scholar
High Temperature-Mediated Adaptations in Plant Architecture Require the bHLH Transcription Factor PIF4
Current Biology
19
:408–413
https://doi.org/10.1016/j.cub.2009.01.046
Google Scholar
A Comparative Transcriptomic Analysis Reveals Conserved Features of Stem Cell Pluripotency in Planarians and Mammals
Stem Cells
30
:1734–1745
https://doi.org/10.1002/stem.1144
Google Scholar
Heat Stress Phenotypes of Arabidopsis Mutants Implicate Multiple Signaling Pathways in the Acquisition of Thermotolerance
Plant Physiology
138
:882–897
https://doi.org/10.1104/pp.105.062257
Google Scholar
Photoperiodic regulation of the C-repeat binding factor (CBF) cold acclimation pathway and freezing tolerance in <em>Arabidopsis thaliana</em>
Proceedings of the National Academy of Sciences
109
:15054–15059
https://doi.org/10.1073/pnas.1211295109
Google Scholar
PIFs: Systems Integrators in Plant Development
The Plant Cell
26
:56–78
https://doi.org/10.1105/tpc.113.120857
Google Scholar
Multiple Phytochrome-Interacting bHLH Transcription Factors Repress Premature Seedling Photomorphogenesis in Darkness
Current Biology
18
:1815–1823
https://doi.org/10.1016/j.cub.2008.10.058
Google Scholar
PIFs: pivotal components in a cellular signaling hub
Trends in Plant Science
16
:19–28
https://doi.org/10.1016/j.tplants.2010.08.003
Google Scholar
Definition of Early Transcriptional Circuitry Involved in Light-Induced Reversal of PIF-Imposed Repression of Photomorphogenesis in Young <em>Arabidopsis</em> Seedlings
The Plant Cell
21
:3535–3553
https://doi.org/10.1105/tpc.109.070672
Google Scholar
Molecular mechanisms governing plant responses to high temperatures
Journal of Integrative Plant Biology
60
:757–779
https://doi.org/10.1111/jipb.12701
Google Scholar
PHYTOCHROME INTERACTING FACTORS PIF4 and PIF5 promote heat stress induced leaf senescence in Arabidopsis
Journal of Experimental Botany
72
:4577–4589
https://doi.org/10.1093/jxb/erab158
Google Scholar
The transcription factors PIF4 and PIF5 interact with WRINKLED1 to modulate fatty acid biosynthesis during seed maturation
Plant Physiology
197
:kiaf141
https://doi.org/10.1093/plphys/kiaf141
Google Scholar
Phytochrome-interacting factors PIF4 and PIF5 negatively regulate anthocyanin biosynthesis under red light in Arabidopsis seedlings
Plant Science
238
:64–72
https://doi.org/10.1016/j.plantsci.2015.06.001
Google Scholar
Phytochrome-mediated inhibition of shade avoidance involves degradation of growth-promoting bHLH transcription factors
The Plant Journal
53
:312–323
https://doi.org/10.1111/j.1365-313X.2007.03341.x
Google Scholar
Plant Development: PIF4 Integrates Diverse Environmental Signals
Current Biology
19
:R265–R266
https://doi.org/10.1016/j.cub.2009.01.051
Google Scholar
Etiolated Hypocotyls: A New System to Study the Impact of Abiotic StressAbiotic stresses on Cell Expansion - Environmental Responses
In:
Duque P
Szakonyi D
, editors.
Plants: Methods and Protocols
Springer US
pp. 195–205
https://doi.org/10.1007/978-1-0716-2297-1_13
Google Scholar
Alternative splicing landscapes in Arabidopsis thaliana across tissues and stress conditions highlight major functional differences with animals
Genome biology
22
:35
https://doi.org/10.1186/s13059-020-02258-y
Google Scholar
The cell biology of tetrapyrroles: a life and death struggle
Trends in Plant Science
15
:488–498
https://doi.org/10.1016/j.tplants.2010.05.012
Google Scholar
A Recently Evolved Alternative Splice Site in the <em>BRANCHED1a</em> Gene Controls Potato Plant Architecture
Current Biology
25
:1799–1809
https://doi.org/10.1016/j.cub.2015.05.053
Google Scholar
Expanding Roles of PIFs in Signal Integration from Multiple Processes
Molecular plant
10
:1035–1046
https://doi.org/10.1016/j.molp.2017.07.002
Google Scholar
A role for an alternative splice variant of PIF6 in the control of Arabidopsis primary seed dormancy
Plant Molecular Biology
73
:89–95
https://doi.org/10.1007/s11103-009-9571-1
Google Scholar
Combinatorial Complexity in a Transcriptionally Centered Signaling Hub in Arabidopsis
Molecular Plant
7
:1598–1618
https://doi.org/10.1093/mp/ssu087
Google Scholar
Phytochromes and Phytochrome Interacting Factors
Plant Physiology
176
:1025–1038
https://doi.org/10.1104/pp.17.01384
Google Scholar
Molecular and genetic control of plant thermomorphogenesis
Nature Plants
2
:15190
https://doi.org/10.1038/nplants.2015.190
Google Scholar
Evolution of Chlorophyll Biosynthesis – The Challenge to Survive Photooxidation
Cell
86
:703–705
https://doi.org/10.1016/S0092-8674(00)80144-0
Google Scholar
Phytochrome-interacting transcription factors PIF4 and PIF5 induce leaf senescence in Arabidopsis
Nature Communications
5
:4636
https://doi.org/10.1038/ncomms5636
Google Scholar
Functional profiling identifies genes involved in organ-specific branches of the PIF3 regulatory network in Arabidopsis
The Plant Cell
23
:3974–3991
https://doi.org/10.1105/tpc.111.088161
Google Scholar
Two splice variants of the IDD14 transcription factor competitively form nonfunctional heterodimers which may regulate starch metabolism
Nature Communications
2
:303
https://doi.org/10.1038/ncomms1303
Google Scholar
Phytochromes promote seedling light responses by inhibiting four negatively-acting phytochrome-interacting factors
Proceedings of the National Academy of Sciences
106
:7660–7665
https://doi.org/10.1073/pnas.0812219106
Google Scholar
PIF3 regulates anthocyanin biosynthesis in an HY5-dependent manner with both factors directly binding anthocyanin biosynthetic gene promoters in Arabidopsis
The Plant Journal
49
:981–994
https://doi.org/10.1111/j.1365-313X.2006.03021.x
Google Scholar
Overexpression of light-dependent PORA or PORB in plants depleted of endogenous POR by far-red light enhances seedling survival in white light and protects against photooxidative damage
The Plant Journal
12
:649–658
https://doi.org/10.1046/j.1365-313X.1997.00649.x
Google Scholar
Arabidopsis SAURs are critical for differential light regulation of the development of various organs
Proceedings of the National Academy of Sciences of the United States of America
113
:6071–6076
https://doi.org/10.1073/pnas.1604782113
Google Scholar
An atlas of alternative splicing profiles and functional associations reveals new regulatory programs and genes that simultaneously express multiple major isoforms
Genome Research
27
:1759–1768
Google Scholar
A Simple Method for Quantification of Protochlorophyllide in Etiolated Arabidopsis Seedlings BT -
In:
Hiltbrunner A
, editors.
Phytochromes: Methods and Protocols
New York
:
Springer
pp. 169–177
https://doi.org/10.1007/978-1-4939-9612-4_14
Google Scholar
The Arabidopsis basic/helix-loop-helix transcription factor family
The Plant Cell
15
:1749–1770
https://doi.org/10.1105/tpc.013839
Google Scholar
Salt-responsive SSN1 condensation in nucleus facilitates PIF4 degradation to regulate Arabidopsis salt tolerance
The Plant Journal
123
:e70389
https://doi.org/10.1111/tpj.70389
Google Scholar
PIF4 Promotes Expression of HSFA2 to Enhance Basal Thermotolerance in Arabidopsis
International Journal of Molecular Sciences
https://doi.org/10.3390/ijms23116017
Google Scholar
Phytochrome interacting factor proteins regulate cytokinesis in Arabidopsis
Cell Reports
35
:109095
https://doi.org/10.1016/j.celrep.2021.109095
Google Scholar
RNA-seq experiment of wild-type (WT) and pif4 mutant seedlings grown for 3 days in darkness and exposed or not to heat stress for 3 hours.
NCBI Gene Expression Omnibus
ID GSE200247
https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE200247
Article and author information
Author information
Cite all versions
You can cite all versions using the DOI
https://doi.org/
10.7554/eLife.110146
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Niño-González et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
0
downloads
0
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.

