model of RVCL-S demonstrates age dependent disease progression

专属客服号

微信订阅号
大数据治理
全面提升数据价值
赋能业务提质增效
Abstract
Retinal vasculopathy with cerebral leukoencephalopathy and systemic manifestations (RVCL-S) is a disease that causes deterioration of small vessels, affecting various organs: eyes, brain, liver, and others. The RVCL-S carriers have lower life expectancy. There is no cure available to date. The disease has been linked to mutations in TREX1 gene disrupting its cytoplasmic localization. To facilitate the disease mechanism investigation, we employed model organism
D. melanogaster,
identified human TREX1 ortholog
cg3165
, and confirmed its vital significance to flies. Then, we expressed human TREX1 and its mutant form TREX1 V235Gfs in flies and used optical coherence microscopy (OCM) to monitor the dynamics of flies’ vascular system. We detected the relapse of fly dorsal vessel, movement impairment, and reduced longevity in TREX1 V235Gfs-expressing transgenic animals. Vascular deterioration and shorter life span recapitulate the RVCL-S manifestations in humans. We have established a robust quantitative
Drosophila
RVCL-S phenotypic system that can potentially serve as a screening platform for drug discovery and drug targets identification.
Introduction
Retinal vasculopathy with cerebral leukoencephalopathy and systemic manifestations (RVCL-S, RVCL) represents a very special case among rare diseases. It is classified as an ultra-rare disease since it was detected in approximately 30 unrelated families from different countries across the world (
1
). The disease manifests in highly vascularized tissues including the central nervous system (CNS), retina, liver, kidney (
2
). The observed symptoms appear between 35 to 50 years of age. Vision impairment, MRI detected brain abnormalities, proteinuria, and liver disease are among the most common symptoms. The severity of the disease correlates with age, leading to premature death (
2
). Proper diagnosis can be established only based on molecular DNA analyses, which is not a common approach taken by a family physician. Most likely, RVCL-S is under-diagnosed and has much broader distribution among the population (
3
).
A hereditary syndrome resulting in brain pseudotumors and retinal capillary abnormalities (cerebroretinal vasculopathy, CRV) was first reported and described in 1988 (
4
). Later, hereditary autosomal dominant vascular retinopathy (HVR), migraine, and Raynaud’s syndrome cases were studied in a large Dutch family (289 family members); the results indicated vascular etiology of this disorder (
5
). Clinical cases of hereditary endotheliopathy with retinopathy, nephropathy, and stroke (HERNS) were observed in Chinese family (
6
). HVR, CRV, and HERNS phenotypes were all linked to the same chromosomal region 3p21.1-p21.3 (
7
). In 2007 these illnesses were designated as retinal vasculopathy with cerebral leukodystrophy (RVCL) and linked to mutations in TREX1 gene resulting in protein C-terminal truncations (
8
,
9
). These mutations are dominant and result in 100% disease penetrance (
8
). RVCL is not associated with elevated levels of Type I interferons (IFNs) (
10
). Development of the appropriate therapy remains very challenging, despite the availability of information related to the TREX1 mutations mapping and the protein function. There are no financial incentives for pharmaceutical companies to invest in large scale drug discovery due to the high cost and a small number of RVCL-S patients. One of the directions for RVCL-S treatment development is drug re-purposing, this approach can reduce the time and associated cost. First clinical trial utilizing Aclarubicin, a component of anti-cancer drug cocktails used in China and Japan, to treat RVCL patients started in 2016 (ClinicalTrials.gov, NCT02723448). However, no benefits were observed and the trial did not advance to Phase II (
11
). Recently, Crizanlizumab, approved for sickle cell anemia treatment, was employed to treat RVCL-S patients in a trial showing a potential to slow the disease progression (
12
).
Using model organisms to create human disease models has been proven to be an indispensable approach to decipher the disease mechanism on molecular and physiological levels and facilitate the treatment development. Introduction of RVCL-S associated mutation mimicking human TREX1 V235G frameshift into a mouse Trex1 gene resulted in increased mortality and vascular phenotypes in homozygous mice; however, not all pathological features of the disease were detected (
13
). Studies of TREX1 function on molecular level using cell culture approaches revealed TREX1 mediated DNA damage and subsequent senescence induction caused by a nuclear envelope rupture (
14
). This may occur naturally in highly mechanically solicited tissues (muscles, etc) due to aging, and also in crowded tissues,
aka
tumors. Chauvin et al. (2024) utilized sophisticated multi-model approach (
Drosophila
, mouse, and cells) to demonstrate the role of RVCL-S causing TREX1protein in accumulation of DNA breaks, cellular senescence induction and loss of specific cell types (
11
). Regardless of a significant progress in understanding the link between TREX1 mutations and RVCL symptoms, there is still a need for a robust disease model to perform high throughput drug screenings; for example, FDA approved compound libraries assessment could help to prioritize the drug candidates.
Drosophila melanogaster
(fruit fly) is an invertebrate model organism with well-studied physiology, behavior, sequenced genome, and availability of sophisticated genetic and biochemical tools. Fruit flies possess significant gene conservation with humans and have been successfully used to create human disease models, including rare disease models (
15
,
16
) (
17
). RVCL-S linked TREX1 gene has an ortholog in
D. melanogaster
,
cg3165,
based on the computational predictions (
18
).
cg3165
is expressed at all developmental stages, as demonstrated by RNA-Seq data (SI Fig. 1)(
19
–
21
).
cg3165
and human TREX1 belong to 3’-5’ exonuclease, DnaQ-like subfamily. Humans have 2 closely related genes, TREX1 and TREX2, their catalytic domains are 40% identical, but TREX2 does not have extended C-terminal region (
22
). TREX2 has nuclear localization and plays a role in genome stability (
23
). The C-terminal region of TREX1 contains the transmembrane domain (TM) anchoring it to perinuclear endoplasmic reticulum (
24
,
25
). TREX1 C-terminal truncations lose their cytoplasmic localization and enter the nucleus causing DNA damage and leading to RVCL-S manifestations (
11
).
cg3165
has significant similarity to TREX1 and TREX2 in N-terminal conserved exonuclease domain, but Alliance of Genome Resources gives higher score to human TREX1 to be an ortholog of
D. melanogaster cg3165
(
18
).
cg3165
does not have a TM domain (
25
), likely using different mechanisms to separate nuclear and cytoplasmic activities.
Drosophila
is the only invertebrate model that has functional and genetically trackable cardiovascular system (
26
). Flies have open circulatory system with a dorsal vessel spanning along the anterior-posterior axis (
27
). We focused our research on creating RVCL-S disease model mirroring genetic aberrations and looking for the phenotypic changes in vascular and other relevant organ systems. Robust phenotypes would be used for creating
Drosophila
based RVCL-S screening platform. Previously, we applied optical coherence microscopy (OCM) for non-invasive,
in vivo
visualization of functioning
D. melanogaster
cardiovascular system, producing images with a micron scale resolution (
28
–
32
). The imaging data were efficiently processed using deep-learning-based neural network models to create masks of vessel cross section area and extract relevant cardiovascular functional parameters (
33
–
36
).
Here, we report the development of an RVCL-S model in
D. melanogaster
. We confirmed the implication of human TREX1 fly ortholog,
cg3165
, in dorsal vessel maintenance, neuromotor regulation, and lifespan prolongation. We have generated a transgenic system, enabling expression of human full length TREX1 and RVCL-S associated C-terminus truncated TREX1 V235G fs in flies. The obtained results have demonstrated the detrimental effects of TREX1 V235G fs on dorsal vessel parameters, neuromotor functions and a lifespan. These instruments can be utilized in search efforts for therapeutic agents to counteract the RVCL-S disease progression.
Results
Genetic design of RVCL-S model in
D. melanogaster
We have identified
Drosophila melanogaster cg3165
as an ortholog of human TREX1 using BLAST and other bioinformatic instruments and searching the genomic and proteomic databases (FlyBase, KEGG, etc.).
cg3165
gene structure is shown in
Fig. 1A
. The first step in generation of RVCL model was aimed to determine the vital significance of
cg3165
gene for
D. melanogaster
. We performed RNAi mediated CG3165 depletion utilizing UAS/GAL4 genetic technique (
37
) (
Table 1
). Ubiquitous depletion (
Act5C> cg3165
RNAi
) resulted in reduced longevity (SI Fig. 2A) and flies’ impaired locomotor behavior (SI Fig. 2C), consistent only in males. Cardiac vessel specific depletion in
Hand>cg3165
RNAi
animals did not result in longevity reduction (SI Fig. 2A, B). These initial results, however, suggested the importance of
cg3165
for the flies’ health. Next, we designed a series of transgenic lines with various
D. melanogaster cg3165
and human TREX1 content in order to optimize and establish a robust genetic system representing the RVCL-S disease state. We have generated several genetic combinations, starting from complete removal of fly CG3165 by CRISPR. Though the complete removal of 3’-5’-DNA exonuclease coded by
cg3165
is rather attributable to Aicardi-Goutieres syndrome (
38
), and is not relevant to the RVCL-S caused by the active enzyme mis-localization, the results obtained with the deletion line were used as a starting point to assess the phenotypic impacts from human TREX1 transgenes. In subsequent steps we gradually added back a copy of
cg3165
gene and introduced transgenic human TREX1 variants. hTREX1 transgenes expression levels were regulated by different GAL4 drivers; details are summarized in
Table 1
.
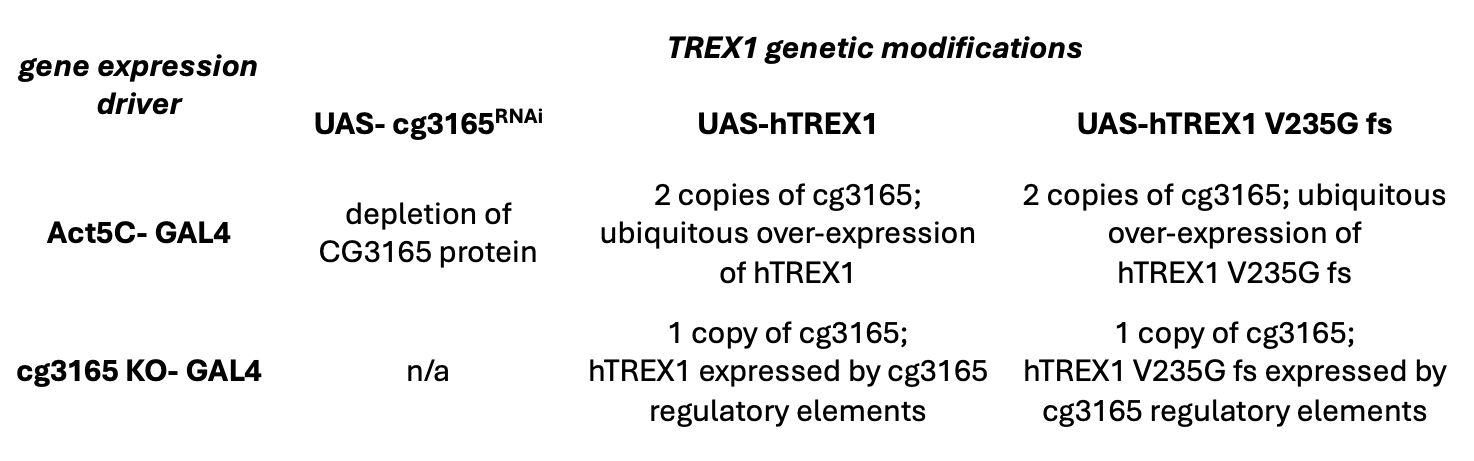
Genetic configuration UAS/GAL4 system for RNAi mediated depletion of
cg3165
and expression of human TREX1 transgenes in
D. melanogaster
Females from GAL4 driver lines listed in 1
st
column were crossed to males shown in the upper row.
The resulting copy numbers of fly
cg3165
and human TREX1 content of the progeny is described.
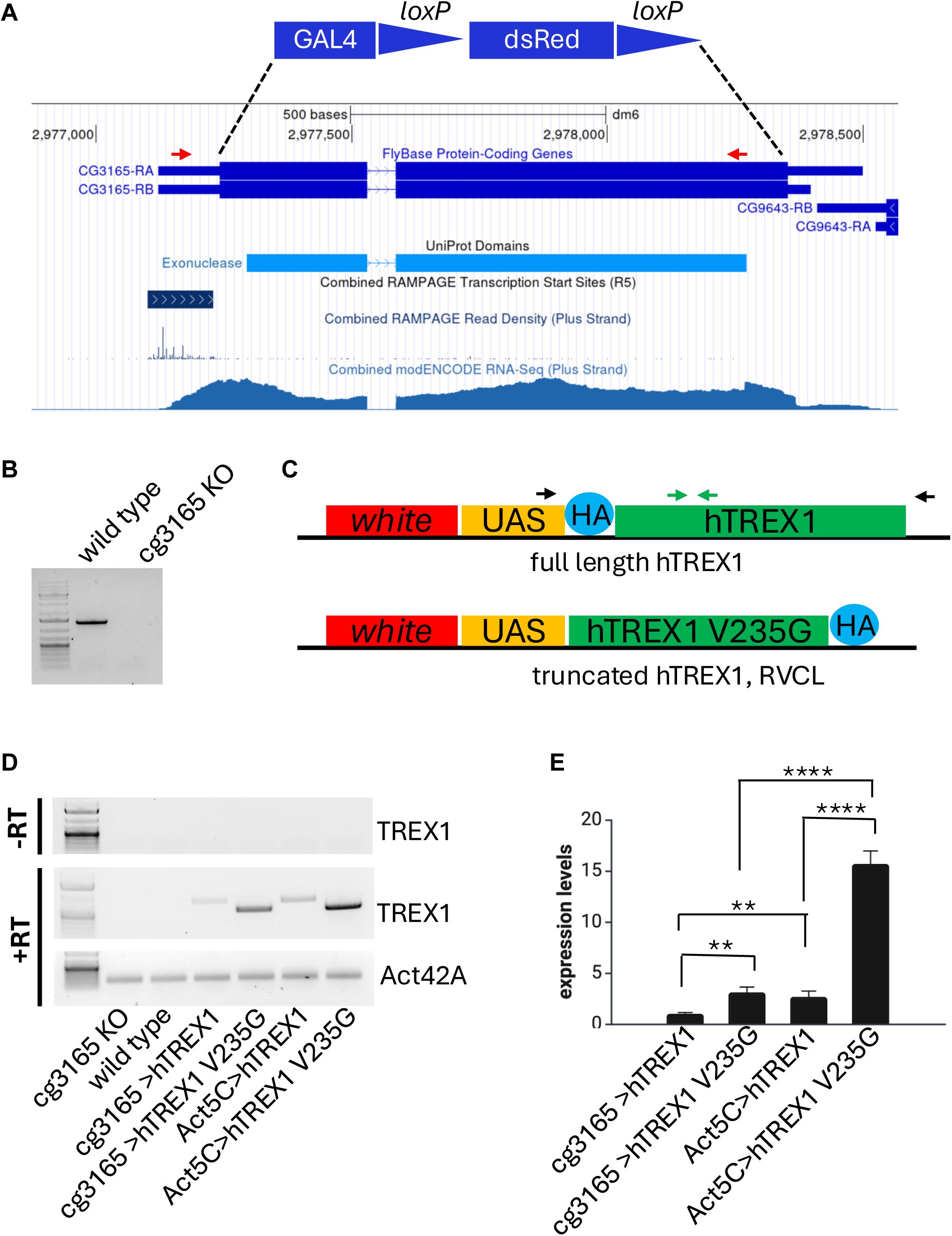
Schematic presentation of generated transgenic constructs and their confirmation after genomic integration and activation.
(A) Genomic region containing
cg3165
gene shown in GEP UCSC browser snapshot with selected tracks indicating protein domain, regulatory regions, and RNA-seq coverage.
cg3165
CDS shown as thick blue blocks; it was replaced by GAL4-lox-dsRed-lox sequence shown between dashed lines (functional elements are not in scale). Red arrows indicate PCR primers used to confirm
cg3165
CDS removal. (B) PCR results confirm the removal of CDS fragment. (C) Transgenic constructs used for establishment of UAS-hTREX1 and UAS-hTREX1 V235G fs
Drosophila
stocks. Black arrows indicate RT-PCR primers. Primers used for quantitative RT-PCR are shown in green. (D) RT-PCR results confirm hTREX1 and hTREX1 V235G fs expression. Genotypes are shown below the gel images.
Act42A
gene was used as a reference. Upper panel represents ‘no reverse transcription’ (-RT) control. First lane is the 1 kb Plus DNA Ladder. Primers’ targets are indicated on the right. (E) Quantitative RT-PCR results demonstrate higher expression levels of both TREX1 transgenes when driven by ubiquitous Act5C-GAL4 driver, compared to cg3165-GAL4 driver and show elevated transcription levels of hTREX1 V235G fs compared to hTREX1. Expression levels were normalized to
Act42A
. Bar graphs represent means with SD; ** p< 0.01; **** p< 0.0001. This panel was created using
BioRender.com
.
We have generated a
cg3165 KO-GAL4
strain by knocking out
cg3165
protein coding DNA sequence (CDS) and knocking-in a GAL4 activator sequence. This line plays a dual role: it represents a null
cg3165
mutant (
Fig. 1A-B
), and, at the same time, it serves as a GAL4 driver regulated by the remaining 5’UTR of
cg3165
gene (
Fig. 1A
, RAMPAGE evidence track)(
39
,
40
). We utilized this GAL4 driver to activate human TREX1 transgenes in
D. melanogaster
presumably natural spatiotemporal pattern. Human TREX1 transgenes were assembled as following: full length TREX1 or truncated TREX1 G235V fs DNA sequences were placed under the control of UAS element in
Drosophila
transformation vector. Both transgenic constructs were incorporated into
D. melanogaster
genome at
attP2
docking site allowing high expression levels (
41
) when activated by GAL4 drivers (
Fig. 1C
, Table 1).
cg3165-GAL4 or Act5C-GAL4
females were crossed to
UAS
-
hTREX1
and
UAS-hTREX1 V235G fs
males and produced viable progeny. We confirmed the expression of
UAS
-
hTREX1
and
UAS-hTREX1 V235G fs
in adult flies (
Fig. 1D
) and evaluated the RNA levels of both transgenes controlled by cg3165-GAL4 and ubiquitous Act5C-GAL4 (
Fig. 1E
). As expected, we observed higher transcript levels for Act5C-GAL4 driven transgenes compared to cg3165-GAL4 driven ones. We also noticed strikingly higher expression of RVCL linked
hTREX1 V235G fs
versus normal
hTREX1
under control of both drivers (
Fig. 1E
). However, we could not detect a truncated hTREX1 V235G fs protein neither with anti-HA (SI Fig. 3A) nor with anti-TREX1(SI Fig. 3B) antibodies by Western blots, where full length hTREX1 produced a band of expected size and intensity (
Act5C> TREX1
is brighter that
cg3165> TREX1
) (SI Fig. 3A-B); these results may reflect the truncated protein instability.
CG3165 knock-out affects the fly dorsal vessel physiology
In RVCL-S patients with diverse symptoms and manifestations, small blood vessels are affected comprehensively. We set up a goal to characterize the parameters change of the only fruit fly’s vessel in response to the presence of RVCL linked human TREX1. To achieve this, we performed a series of genetic manipulations and assessed our disease model at different building steps. We started with the complete removal of fly CG3165. Young seven day old
cg3165 KO-GAL4
adults’ dorsal vessels were subjected to OCM imaging, males and females were imaged separately (
32
). Imaging data quantitation suggests that the physiology of the fly dorsal vessel was changed (
Fig. 2
, SI Video 1-2, SI Fig. 4). We determined several parameters including heart rates (HR), end diastolic area (EDA), end systolic area (ESA), fractional shortening (FS), and arrhythmicity index (AI).
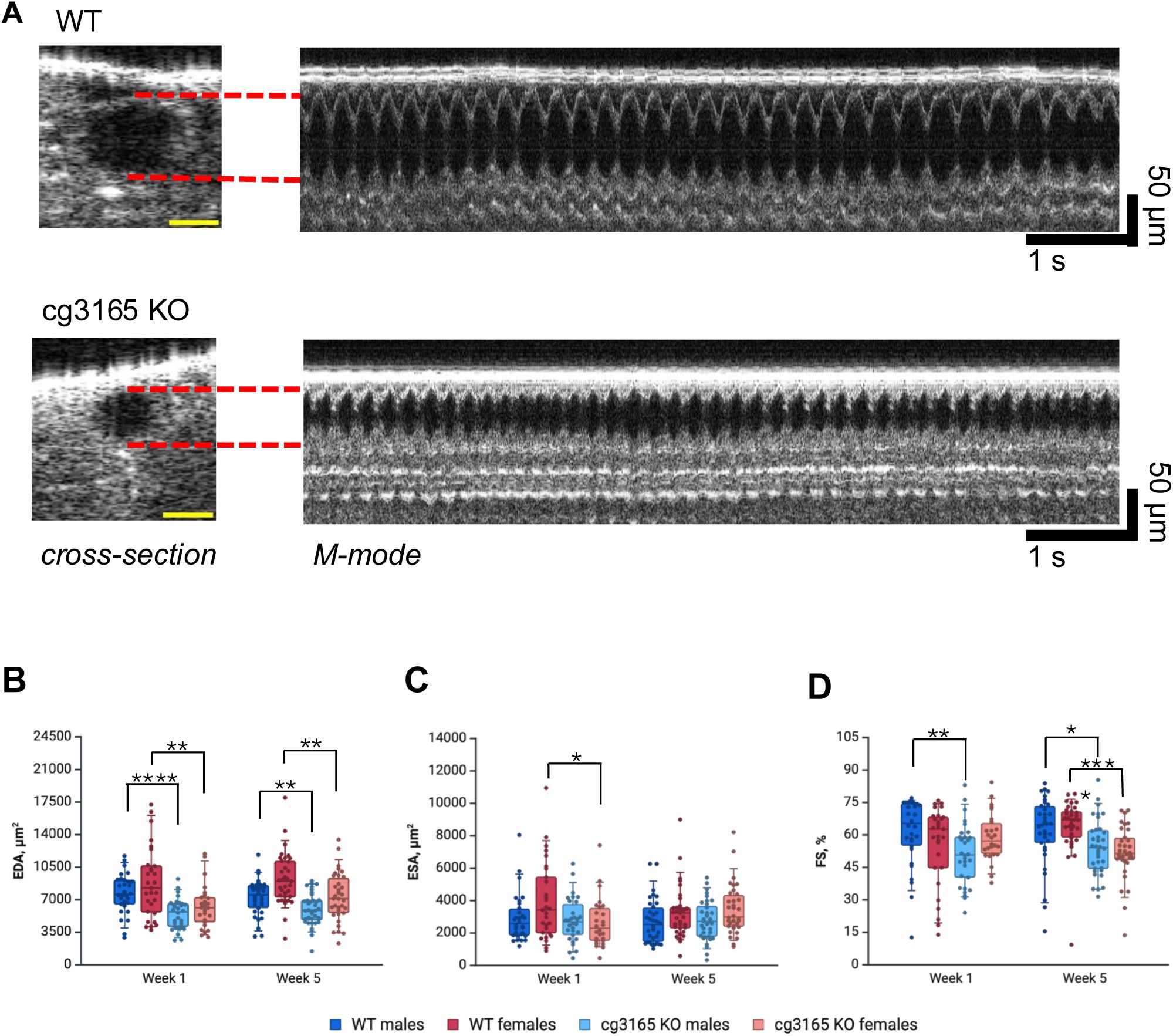
cg3165 knock out affects
D. melanogaster
dorsal vessel functional parameters.
(A) Cross-section and M-mode OCM images of WT and cg3165 KO-GAL4 flies (7 days old males shown). Yellow scale bar is 50 um. (B) End diastolic area (EDA) measurements. (C) End systolic area (ESA)measurements. (D) Fractional shortening (FS) parameters. One week and 5 weeks old flies shown in panels B-D; males are shown as blue shade boxes, females shown as red shade boxes. 1 week old sample sizes: WT males n=30, WT females n=29; cg3165 KO-GAL4 WT males n=30, KO-GAL4 WT females n=26. 5 weeks old sample sizes: WT males n=34, WT females n=35; cg3165 KO-GAL4 males n=36, cg3165 KO-GAL4 females n=34. Statistical significance shown as black brackets. * - p< 0.05; ** - p< 0.01; *** - p< 0.001; **** - p< 0.0001. Panels B and D were created using
BioRender.com
.
cg3165
KO-GAL4 flies demonstrated vessel’s impaired ability to dilate as judged by the EDA measurements; both males and females were affected (
Fig. 2B
). The average EDA in 1-week-old WT males is 7.7 ± 0.4 x 10
3
μm
2
vs 5.5 ± 0.3 x 10
3
μm
2
in
cg3165 KO-GAL4
(both n =30, p <0.0001); the females’ average EDA is 8.8 ± 0.7 x 10
3
μm
2
vs 6.3 ± 0.5 x 10
3
μm
2
respectively (WT n = 29;
cg3165 KO-GAL4
n = 30; p < 0.01). This phenotype was observed in young flies (1-week-old) as well as in aged flies (5-week-old). The average EDA in 5-week-old WT males is 7.2 ± 0.3 x 10
3
μm
2
vs 6.0 ± 0.3 x 10
3
μm
2
in
cg3165 KO-GAL4
(WT n = 34;
cg3165 KO-GAL4
n = 36; p < 0.01); the females’ average EDA is 9.2 ± 0.5 x 10
3
μm
2
vs 7.3 ± 0.4 x 10
3
μm
2
respectively (WT n = 35;
cg3165 KO-GAL4
n = 34; p < 0.01).
Reduction of the vessel diameter during maximum contraction, ESA, in
cg3165
KO flies was observed in 1-week-old females (
Fig. 2C
); WT 1 week old females ESA average is 4.0 ± 0.4 x 10
3
μm
2
and
cg3165 KO-GAL4
is 2.7 ± 0.3 x 10
3
μm
2
(WT n = 29;
cg3165 KO-GAL4
n = 26; p < 0.05). One week old males have comparable ESA measurements; WT average is 2.9 ± 0.3 x 10
3
μm
2
and cg3165 KO-GAL4 is 2.8 ± 0.3 x 10
3
μm
2
(WT n = 30;
cg3165 KO-GAL4
n = 30; p > 0.05). Five-week-old males and females ESA were not affected by the removal of CG3165 (
Fig. 2C
).
Fractional shortening (FS) parameter reflects
Drosophila
dorsal vessel contractility; it measures the percentage difference between the diastolic and systolic states of the fly vessel. We observed impaired contracting ability in 1 week old
cg3165
null males, FS reduction in females was not statistically significant. However, in older flies, both males and females have demonstrated impaired contractility. In 1-week-old WT males, the average FS is 61.5 ± 2.9 % and FS of
cg3165 KO-GAL4
1-week-old males is significantly lower, 51.0 ± 2.6 % (both genotypes n = 30, p < 0.01). FS of WT and
cg3165 KO-GAL4
1 week old females is not significantly different, 55.5 ± 3.4 % and 59.0% ± 2.2 % respectively (WT n = 29;
cg3165 KO-GAL4
n = 26; p >0.05,
Fig. 2D
). The average FS in 5-week-old WT males is 62.0 ± 2.7 % vs 54.3 ± 2.1 % in
cg3165 KO-GAL4
(WT n = 34;
cg3165 KO-GAL4
n = 36; p < 0.05); the females’ average FS is 63.9 ± 2.1% vs 51.9 ± 2.1%, respectively (WT n = 35;
cg3165 KO-GAL4
n = 34; p < 0.0001,
Fig. 2D
).
Analyzing the other characteristics of fly cardiovascular system, such as heart rates (HR) and arrhythmicity indexes (AI), we detected no significant effects from CG3165 ablation. Of note, the distinct HR were observed between two age groups (1-week-old and 5-week-old flies). In
Drosophila,
HR tends to reduce with age (
42
,
43
); we perceived this in WT and
cg3165
KO-GAL4 flies comparing 1-week-old with 5-week-old males and females, further validating our research tools (SI Fig. 4A). The rhythmicity (AI) displays of
cg3165 KO-GAL4
and WT flies were similar across both age groups (SI Fig. 4B). These results suggest no involvement of
cg3165
in heart regulation
per se
. However, the functioning pattern of the
cg3165
KO-GAL4 flies cardiac vessel is noticeably different from the WT and some phenotypic manifestations (EDA, FS) appear to be related to vessel ‘rigidity’ and are age sensitive. Because the most robust changes were observed in EDA values across both sex and age groups, we decided to use primarily EDA read-out in our following experiments.
Transgenic hTREX1 rescues the phenotypes caused by
cg3165
knock-out
After we cleared
D.melanogaster
genome of endogenous CG3165 and ascertained the vascular phenotypes, we introduced transgenic human TREX1 or RVCL-S associated TREX1 V235G fs to test the ability of human TREX1 to rescue the vascular phenotypes caused by the
cg3165
knock-out. We crossed
cg3165 KO-GAL4
line carrying 0 copies of
cg3165
with
UAS-hTREX1
or
UAS-hTREX1 V235G fs
line containing hTREX1 transgene and 2 copies of endogenous
cg3165
(SI Fig. 5). Therefore, the resulting
cg3165>TREX1
or
cg3165>TREX1 V235G fs
progeny contains 1 copy of
cg3165
and an activated hTREX1 transgene (Table 1, SI Fig. 5). This balanced genetic design was applied to prevent possible negative effects from an introduction of human protein into fly organism.
To assess the vascular phenotypes, flies were subjected to OCM imaging;
cg3165> hTREX1
(1x
cg3165
, 1x hTREX1) and
cg3165> hTREX1 V235G
(1x
cg3165
, 1x hTREX1 V235G fs) were compared to WT (2x
cg3165
, 0x hTREX1),
cg3165 KO-GAL4
(0x
cg3165,
0x hTREX1), and
cg3165> yw
(1x
cg3165,
0x hTREX1). Adult flies’ dorsal vessel was imaged at day 7 (1 week) and day 35 (5 weeks) post eclosure. The EDA measurements are summarized in
Fig. 3A-B
; SI Video 1-4 contain representative videos. The vessel’s diastolic diameter is increased upon the introduction of a copy of cg3165 and transgenic hTREX1 and TREX1 V235G fs as seen in young males (1-week-old).
cg3165 KO-GAL4
average EDA is 5.5 ± 0.3 x 10
3
μm
2
versus 7.0 ± 0.4 x 10
3
μm
2
in
cg3165> yw
(both n = 30, p < 0.01), 7.5 ± 0.5 x 10
3
μm
2
in
cg3165> hTREX1
(n = 26, p < 0.01), and 7.6 ± 0.5 x 10
3
μm
2
in
cg3165> hTREX1 V235G
(n = 25, p <0.01) (
Fig. 3A
). One-week-old
cg3165> yw
,
cg3165> hTREX1
, and
cg3165> hTREX1 V235G
females also show statistically larger EDA, compared to
cg3165 KO-GAL4
(
Fig.3B
). However,
cg3165> hTREX1 V235G
average EDA (n = 29) is smaller than
cg3165> yw
(n = 30) hemizygous control (8.2 ± 0.4 x 10
3
μm
2
vs 9.9 ± 0.7 x 10
3
μm
2
, p < 0.05,
Fig. 3B
). The aged hTREX1 and hTREX1 V235G transgenic flies (5 weeks after eclosion) demonstrate vascular relapse, as the EDA values of
cg3165 >TREX1
and
cg3165 >TREX1 V235G fs
flies are smaller than in control
cg3165 >yw
; this effect is observed for both males and females. The average EDA in 5-week-old
cg3165> yw
males (n = 35) is 8.7 ± 0.4 x 10
3
μm
2
versus 7.5 ± 0.4 x 10
3
μm
2
in
cg3165> hTREX1
(n = 36), and 7.0 ± 0.4 x 10
3
μm
2
in
cg3165> hTREX1 V235G
(n = 35) (p < 0.05 and p < 0.01, respectively); the 5-week-old females’ average EDA in
cg3165> yw
(n = 36) is 11.4 ± 0.5 x 10
3
μm
2
versus 8.5 ± 0.4 x 10
3
μm
2
in
cg3165> hTREX1
(n = 31), and 9.3 ± 0.5 x 10
3
μm
2
in
cg3165> hTREX1 V235G fs
(n = 34, p< 0.0001 and p<0.01, respectively). However,
cg3165> hTREX1
and
cg3165> hTREX1 V235G fs
EDA remains significantly larger than the EDA of 5-week-old
cg3165 KO-GAL4
flies (
Fig. 3A-B
).
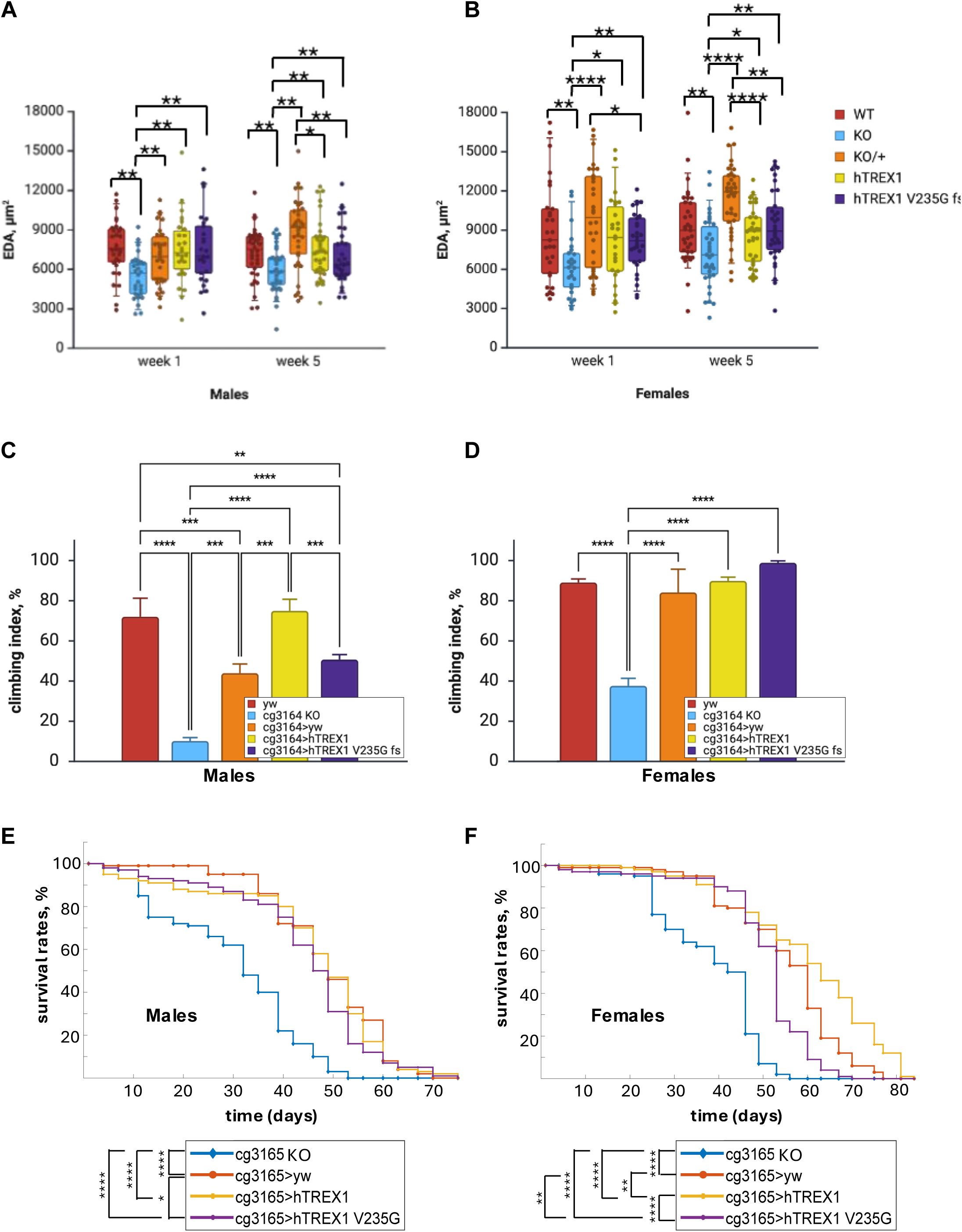
EDA reduction, behavioral impairment, and reduced survival probability observed in CG3165 KO flies are restored by transgenic expression of human TREX1 variants.
(A) One-week-old males’ EDA is significantly larger in
cg3165> yw, cg3165> TREX1, and cg3165> TREX1 V235G fs
than in
cg3165 KO
. Five-week-old males carrying hTREX1 and hTREX1 V235G fs demonstrate reduction of EDA compared to
cg3165> yw
. (B) One-week-old females’ EDA is significantly larger in
cg3165> yw, cg3165> TREX1, and cg3165> TREX1 V235G fs
than in
cg3165 KO
. Five-week-old females carrying hTREX1 and hTREX1 V235G fs demonstrate reduction of EDA compared to
cg3165> yw
. WT-
yw
, KO-
cg3165 KO-GAL4
, KO/+ -
cg3165> yw
;
hTREX1 - cg3165> hTREX1
; hTREX1 V235G fs -
cg3165> TREX1 V235G fs
. 1 week old sample sizes: WT males n=30, KO males n=30, KO/+ males n=30, hTREX1 males n=26, hTREX1 V235G fs males n=25; WT females n=29, KO females n=26, KO/+ females n=30, hTREX1 females n=26, hTREX1 V235G fs females n=29. 5 weeks old sample sizes: WT males n=34, KO males n=36, KO/+ males n=35, hTREX1 males n=36, hTREX1 V235G fs males n=34; WT females n=35, KO females n=34, KO/+ females n=36, hTREX1 females n=31, hTREX1 V235G fs females n=34. (C)
cg3165
knock-out males show impaired locomotor behavior compared to WT (
yw
); hemizygous
cg3165
(
cg3165> yw
) males demonstrate significant improvement. hTREX1 introduction (
cg3165>
hTREX1) results in climbing ability improvement comparable to WT levels. Males carrying RVCL-S associated hTREX1 V235G fs (
cg3165> TREX1 V235G fs
) demonstrate partial climbing ability restoration. (D)
cg3165
knock-out females show impaired climbing compared to WT (
yw
). Normal locomotor behavior is observed in
cg3165> yw, cg3165> TREX1, and cg3165> TREX1 V235G fs
animals. Bar graphs (A-B) show means with SD. (E) Kaplan-Meier survival curves of
cg3165
KO,
cg3165 >yw
,
cg3165 >hTREX1
and
cg3165 > hTREX1 V235G fs
males. Survival rates of
cg3165
knock-out males are significantly reduced. Upon introduction of 1 copy of
cg3165
(
cg3165 >yw
) and transgenic hTREX1 (
cg3165 >hTREX1
), or hTREX1 V235G fs (
cg3165 > hTREX1 V235G fs
) the survival probability is increased. (F) Kaplan-Meier survival curves of
cg3165
KO,
cg3165 >yw
,
cg3165 >hTREX1
and
cg3165 > hTREX1 V235G fs
females. Survival rates of
cg3165
knock-out females are significantly reduced. Upon introduction of 1 copy of
cg3165
(
cg3165 >yw
) and transgenic hTREX1 (
cg3165 >hTREX1
), or hTREX1 V235G fs (
cg3165 > hTREX1 V235G fs
) the survival probability is increased. hTREX1 introduction has the strongest impact on longevity; hTREX1 V235G fs effect is the weakest. Statistical significance shown as black brackets.: * - p< 0.05; ** - p< 0.01; *** - p< 0.001; **** - p< 0.0001. Panels A–D were created using
BioRender.com
.
Analyzing the EDA phenotypes in current hTREX1 expression system, we observed the rescue by the hTREX1 and hTREX1 V235G fs of a vascular phenotype caused by CG3165 knock-out, though we did not detect the significant differences between the full length hTREX1 and hTREX1 V235G fs efficiency. Comparing 1-week and 5-week post eclosure groups we noticed aging-related detrimental effects caused by hTREX1 V235G fs, and also by a full length hTREX1 in males and females (
Fig. 3A-B
).
RVCL affects multiple organ systems and has neurological manifestations including brain dysfunction (
9
). Fruit flies’ survival greatly depends on their motor functions; this is achieved by tight coordination between the CNS processing external signals and sending instructions, motor neurons, and muscles performing certain actions. Disruption of these processes results in motor defects (
44
). We performed a climbing assay to test the ability of TREX1 mutant to maintain a normal behavioral pattern of negative geotaxis. We observed that the
cg3165
knock-out resulted in significantly impaired climbing ability of
cg3165 KO-GAL4
flies; climbing index dropped from WT 72.0 ± 9.2% to 10.1 ± 1.7% (p < 0.0001) in
cg3165 KO-GAL4
males; from 89.1 ± 1.8% to 37.6 ± 3.7% (p < 0.0001) in females (
Fig. 3C-D
), suggesting that the
cg3165
gene plays a broad role affecting the neurological networks and/or muscle tissue condition. The climbing ability has significantly improved in hemizygous
cg3165 >yw
(climbing index is 44.0 ± 4.5% for males (p < 0.001), 84.1 ± 11.5% (p < 0.0001) for females) and in flies expressing both hTREX1 variants:
cg3165 >hTREX1
(75.0 ± 5.7%, p < 0.0001 for males; 89.9 ± 1.8%, p < 0.0001 for females),
cg3165 >hTREX1 V235G fs
flies (50.7 ± 2.5% (p < 0.0001) for males; 98.9 ± 1.0% (p < 0.0001) for females) (
Fig. 3C-D
). In particular, males show remarkable gene dose sensitivity in restoration of locomotor behavioral patterns. Adding hTREX1 transgenic copy significantly increased the climbing index compared to
cg3165 KO-GAL4 and cg3165 >yw,
while the effect from RVCL-S associated hTREX1 V235G fs was significantly less compared to hTREX1 carrying flies (
Fig. 3C
).
Lifespan is a robust indicator of aging rates in fly population. We performed the longevity study (25C, 70 % humidity) to determine the effects of
cg3165
knock-out and introduction of human transgenic copies of full length or truncated hTREX1 forms. Our initial experiments implied that CG3165 depletion reduced the lifespan, but only in males (SI Fig. 2A). Complete CG3165 removal significantly reduces the life span of
cg3165 KO-GAL4
flies (
Fig. 3E-F
, SI Table 1), males and females, compared to
cg3165 >yw
(p < 0.0001 for males, p < 0.0001 for females)
, cg3165 >hTREX1
(p < 0.0001 for males, p < 0.0001 for females) and
cg3165 > hTREX1 V235G fs
flies (p < 0.0001 for males, p < 0.0001 for females). Survival probability was increased in
cg3165 >hTREX1
females, compared to
cg3165 >yw
control (p < 0.01,
Fig. 3F
, SI Table 1); females carrying RVCL associated hTREX1 V235G fs demonstrate reduced longevity compared to
cg3165 >yw
(p < 0.01), and to
cg3165 >hTREX1
(p < 0.0001).
cg3165 >yw, cg3165 KO-GAL4>hTREX1
and
cg3165 >hTREX1 V235G fs
males show similar lifespan patterns (
Fig. 3E
), close to average
Drosophila melanogaster
male lifespan at 25C (
45
).
Over-expression of human TREX1 transgenes show the age dependent phenotypes
To enhance the phenotypic effects from the ectopic hTREX1 and hTREX1 V235G fs expression we decided to use strong ubiquitous Act5C-GAL4 driver to increase the transgene transcription rates (
Fig. 1E
). We performed crosses between
Act5C-GAL4
females and
UAS-hTREX1
and
UAS-hTREX1 V235G fs
males (Table 1, SI Fig.6). One needs to point out, that these genetic configurations include 2 copies of
cg3165
(SI Fig. 6), not 1, as in
cg3165 >hTREX1 and cg3165 >hTREX1 V235G fs
flies (Table 1, SI Fig.5). Higher
Act5C-GAL4 driven UAS-hTREX1
and
UAS-hTREX1 V235G fs
expression levels compared to
cg3165 >hTREX1 and cg3165 >hTREX1 V235G fs
flies’ levels (
Fig. 1E
) did not lead to an increased lethality during early developmental stages of hTREX1 and hTREX1 V235G fs expressing progeny compared to non-transgenic siblings. Obtained
Act5C >hTREX1 V235G fs
flies’ longevity measurements have shown notably shorter lifespan relative to
Act5C>hTREX1
(p < 0.0001 for males and p < 0.0001 for females), and
Act5C>yw
animals (p < 0.0001 for males and p < 0.001 for females) (
Fig. 4A-B
, SI Table 2). hTREX1 expressing females have demonstrated increased longevity compared to control
Act5C>yw
(p < 0.001) (
Fig. 4B
), while
Act5C >hTREX1
males’ lifespan was similar to the control
Act5C>yw
males (p > 0.05) (
Fig. 4A
). These results clearly demonstrate the negative effects of RVCL-S linked hTREX1 V235G fs on flies’ survival probability.
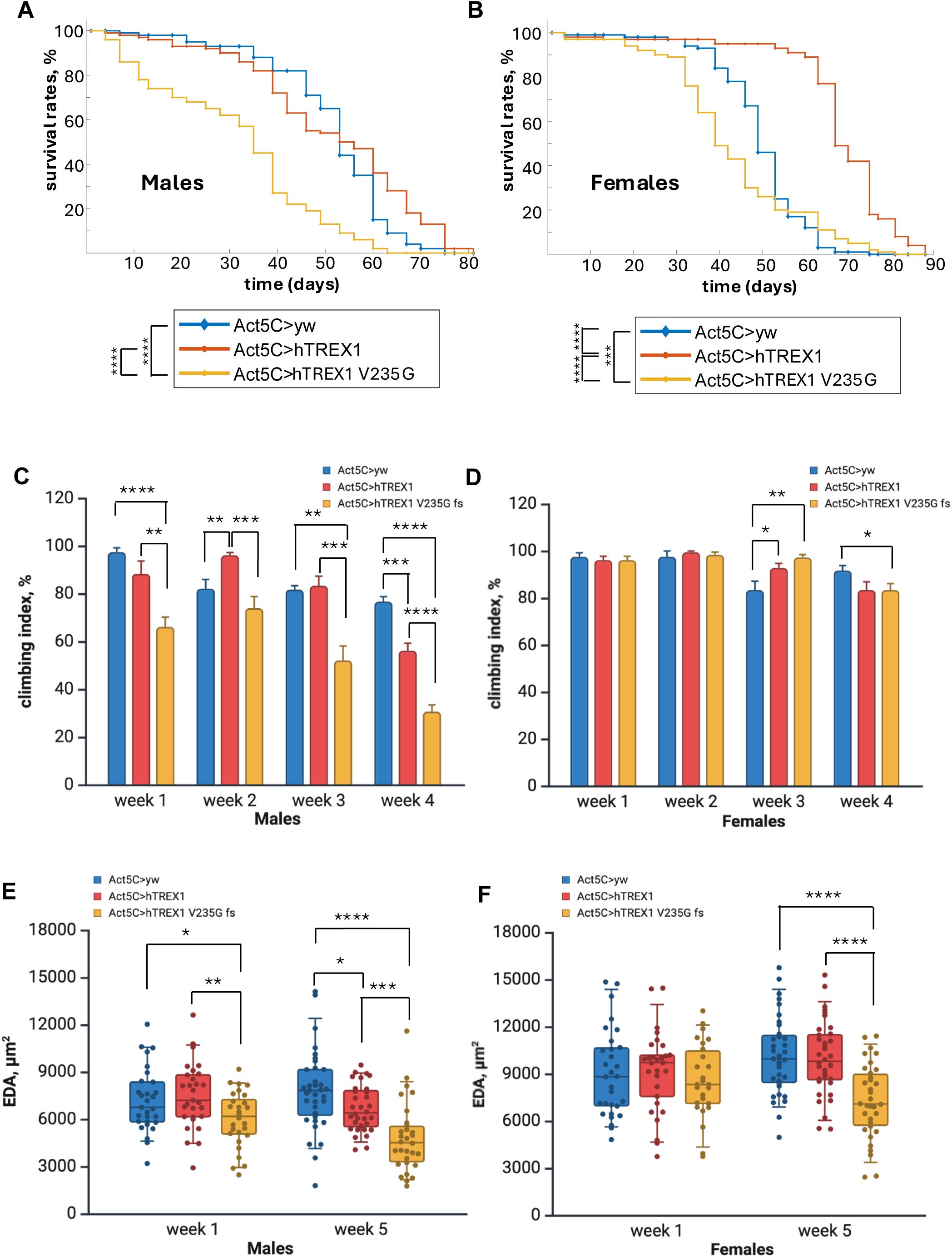
Over-expression of RVCL linked hTREX1 V235G fs reduces survival probability, movement impairment, and decrease of dorsal vessel EDA.
In aged flies, over-expression of ‘healthy’ hTREX1 transgene negatively affects climbing ability and results in smaller EDA. (A) Kaplan-Meier survival curves of
Act5C>yw
(blue line),
Act5C>hTREX1
(orange line) and
Act5C >hTREX1 V235G fs
(yellow line) males. (B) Kaplan-Meier survival curves of
Act5C>yw
(blue line),
Act5C>hTREX1
(orange line) and
Act5C >hTREX1 V235G fs
(yellow line) females. C- Climbing assay results in
Act5C
>
yw
,
Act5C
>
hTREX1
,
Act5C
>
hTREX1 V235G fs
males. D- Climbing assay results in
Act5C
>
yw
,
Act5C
>
hTREX1
,
Act5C
>
hTREX1 V235G fs
females. Bar graphs (C, D) represent means with SEM. (D) End Diastolic Area (EDA) measurements of
Act5C
>
yw
,
Act5C
>
hTREX1
,
Act5C
>
hTREX1 V235G fs
males. (E) End Diastolic Area (EDA) measurements of
Act5C
>
yw
,
Act5C
>
hTREX1
,
Act5C
>
hTREX1 V235G fs
females. 1 week old groups sample sizes:
Act5C
>
yw
males n=29,
Act5C
>
hTREX1
males n=29,
Act5C
>
hTREX1 V235G fs
males n=28; WT females n=30, hTREX1 females n=29,
Act5C
>
hTREX1 V235G fs
females n=26. 5 week old groups sample sizes:
Act5C
>
yw
males n=25,
Act5C
>
hTREX1
males n=32,
Act5C
>
hTREX1 V235G fs
males n=30;
Act5C
>
yw
n=35,
Act5C
>
hTREX1
females n=36,
Act5C
>
hTREX1 V235G fs
females n=34. Statistical significance shown as black brackets. * - p< 0.05; ** - p< 0.01; *** - p< 0.001; **** - p< 0.0001. Panels C–F were created using
BioRender.com
.
Behavioral tests were performed on aging flies, from week 1 to week 4 after eclosion, to assess the effect of mutant hTREX1 V235G fs over-expression (
Fig. 4C-D
). Impaired climbing ability was observed in
Act5C >hTREX1 V235G fs
males starting from the young age, at week 1 and persisted to the end point.
Act5C >hTREX1 V235G fs
females started showing the moving impairment much later, at week 4. The
Act5C>hTREX1
and
Act5C >hTREX1 V235G fs
flies’ responses to climbing challenge were similar to the results observed for 1 week old
cg3165 >hTREX1
and
cg3165 >hTREX1 V235G fs
flies (
Fig. 3C-D
). Males were more sensitive to the detrimental effect of RVCL-S linked hTREX1 mutation. We also noticed the age correlated negative impact from hTREX1 over-expression in both males and females, at week 4 after eclosion, compared to
Act5C>yw
control (
Fig. 4C
).
Considering the physiological impacts of hTREX1 isoforms over-expression, we performed the OCM imaging of flies’ cardiovascular system at week 1 and week 5 after eclosion. The EDA measurements extracted from the processed imaging data are summarized in
Fig. 4E-F
, SI Video 5-7. At week 1, we observed the EDA reduction in
Act5C >hTREX1 V235G fs
(RVCL) males (6.0 ± 0.3 x 10
3
μm
2
, n = 28) relatively to control
Act5C>yw
(7.2 ± 0.4 x 10
3
μm
2
, n = 29, p < 0.05) and
Act5C>hTREX1
(7.5 ± 0.4 x 10
3
μm
2
, n = 29, p < 0.005) (
Fig. 4E
); females average EDA were:
Act5C>yw
- 9.0±0.5 x 10
3
μm
2
(n = 30),
Act5C>hTREX1 -
9.2 ± 0.5 x 10
3
μm
2
(n = 29),
Act5C>hTREX1 V235G fs -
(8.6 ± 0.5 x 10
3
μm
2
, n = 26); the EDA reduction in
Act5C>hTREX1 V235G fs
females is not statistically significant (p>0.05) (
Fig. 4F
). At week 5, we continued to see EDA reduction in males carrying hTREX1 V235G fs (RVCL). The average EDA of
Act5C>hTREX1 V235G fs
5-week-old males are 4.8± 0.4 x 10
3
μm
2
(n = 30). It is significantly smaller than the average EDA of 5-week-old
Act5C>yw
males (7.8 ± 0.4 x 10
3
μm
2
, n = 35, p < 0.0001), and the average EDA of 5-week-old
Act5C>hTREX1
males (6.7 ± 0.3 x 10
3
μm
2
, n = 32, p < 0.001). We also observed the negative effects on EDA in aged
Act5C>hTREX1
males, expressing ‘healthy’ hTREX1. The average EDA of 5-week-old
Act5C>hTREX1
males (6.7 ± 0.3 x 10
3
μm
2
, n = 32) is smaller than the average EDA of 5-week-old
Act5C>yw
males, (7.8 ± 0.4 x 10
3
μm
2
, n = 35, p < 0.05,
Fig. 4E
). The detrimental effect of hTREX1 V235G fs expression became evident in aged females. The average EDA of
Act5C>hTREX1 V235G fs
5-week-old females are 7.3 ± 0.4 x 10
3
μm
2
(n= 34). It is significantly smaller than the average EDA of 5-week- old
Act5C>yw
females
(
10.2 ± 0.4 x 10
3
μm
2
, n = 35, p < 0.0001), and the average EDA of 5-week-old
Act5C>hTREX1
females (10.0 ± 0.4 x 10
3
μm
2
, n = 36, p < 0.0001) (
Fig. 4F
).
Overall, the longevity test, behavioral assays, and OCM imaging results point to the hTREX1 V235G fs as an adverse modulator of physiological processes in adult flies.
Discussion
We took advantage of
Drosophila melanogaster
genetic capability and an innovative imaging technology, OCM, to create an RVCL-S disease model that will facilitate the screening for therapeutics and could be used to study the disease progression. We have identified
D. melanogaster
ortholog of human TREX1 gene linked to the disorder,
cg3165,
and confirmed its significance for the flies’ vital functions. The removal of
cg3165
CDS lead to fly’s cardiovascular system changes comprehensively evaluated by OCM. Dorsal vessel parameter deviations were used as a phenotypic read-out for the RVCL-S fly model building, where the end diastolic area (EDA) proved to be the most robust.
Drosophila
transgenic lines, carrying the full length human TREX1 and RVCL-S associated truncated TREX 1 V235G fs were generated. We focused our efforts on uncovering the distinct phenotypes caused by the full length and the mutant hTREX1. We detected that the flies’ vascular integrity was restored upon the introduction of both human TREX1 transgenes and/or a copy of
cg3165
(
Fig. 3
, SI Fig. 5) compared to
cg3165
null. However, we could not distinguish between the impacts from the hTREX1 and hTREX1 V235G fs at this point, the EDA parameters of
cg3165> hTREX1
and
cg3165> hTREX1 V235G fs
animals were similar.
We extended our assays to behavioral tests based on the rational that RVCL-S affects cerebral functions (
1
). Upon
cg3165
knock-out, flies have demonstrated a significant movement impairment at week 1 after eclosure compared to the WT (
Fig. 3C-D
). Adding a copy of
cg3165
, hTREX1, or hTREX1 V235G fs improved the climbing ability compared to the
cg3165
KO. In males, we observed gradual effects:
cg3165 >yw
demonstrated some improvement; hTREX1 carrying flies’ climbing index was the highest; but hTREX1 V235G fs flies’ climbing ability was significantly lower than of hTREX1 carrying animals. These results suggested strong positive correlation between
Drosophila
neuromotor regulation and transgenic hTREX1 presence and a negative impact from hTREX1 V235G.
In addition to the experiments scrutinizing the organ specific functions related to the RVCL-S, we performed a longevity study of flies with various hTREX1 genetic content (
Fig. 3E-F
).
cg3165
KO animals are homozygous viable, but their lifespan is significantly shorter. Adding back
cg3165
, hTREX1, or hTREX1 V235G fs prolongs the lifespan. The impact is higher in
cg3165> hTREX1
and is less in
cg3165> hTREX1 V235G fs
flies relatively to the
cg3165>yw
control as seen in females. Longevity is not a phenotype that can be used for screening purposes, but it is very reliable method to strengthen the RVCL-S model in
Drosophila.
RVCL-S patients have decreased life expectancy (
2
), and our results with hTREX1 V235G fs carrying flies reflect this disease aspect.
Following the observed trends in our results, we changed the genetic content of the experimental setup; instead of maintaining the gene expression pattern and copy numbers (1 copy of endogenous
cg3165
and a human hTREX1/ hTREX1 V235G fs transgene controlled by 5’UTR cg3165), we utilized ubiquitous
Act5C-GAL4
driver to over-express human TREX1 constructs (SI Fig. 6) anticipating to obtain more robust phenotypes. The longevity evaluation of
Act5C>yw
(control),
Act5C> hTREX1
, and
Act5C> hTREX1 V235G fs
flies re-confirmed the negative impact of hTREX1 V235G fs expression on
Drosophila
survival rates (
Fig. 4A-B
). The locomotor behavior tests performed during week 1 through week 4 demonstrated progressive impairment of climbing ability of hTREX1 V235G carrying males relatively to hTREX1 carrying and control animals (
Fig. 4C
).
The OCM imaging results of
Act5C>yw
,
Act5C> hTREX1
, and
Act5C> hTREX1 V235G fs
clearly demonstrated the detrimental effect of hTREX1 V235G fs on dorsal vessel dilation in 1-week-old males compared to hTREX1 and ‘no transgene’ control (
Fig. 4E
). Considering the late onset of RVCL-S manifestations in humans, in aged (5-week-old) flies, we detected the TREX1 V235G fs facilitated the EDA reduction in both males and females (
Fig.4E-F
).
To build an RVCL-S research model, we devised two
Drosophila
UAS/GAL4 expression systems of human TREX1 and TREX1 V235G fs proteins:
cg3165
and
Act5C
promoter based. The Act5C over-expression approach proved to be more efficient to detect the detrimental effect of RVCL-S linked hTREX1 V235G fs on vascular phenotypes. But the more robust EDA phenotype appearance could also be affected by the fly
cg3165
gene copy numbers in the background (
i.e.
, amount of 3’-5’-DNA exonuclease). McGlasson
et al.
(2025) have recently shown that mono-allelic truncating mutations in TREX1 require intact nuclease activity in order to cause endothelial disease (
46
). The EDA measurements, shown in
Fig. 3A-B
, do not indicate any differences between
cg3165> hTREX1
and
cg3165> hTREX1 V235G fs
animals in any sex or age groups. Meanwhile, the dorsal vessel deterioration (smaller EDA) in
Act5C> hTREX1 V235G fs
is obvious in young and aged males, and aged females, compared to of
Act5C> hTREX1
(
Fig. 4E-F
). The absence of EDA phenotypic differences in the 1
st
case correlates with
cg3165
haplodeficiency (SI Fig. 5) and, therefore, lower level the exonuclease activity. The
Act5C
based expression system includes two copies of
cg3165
(SI Fig. 6) and has higher levels 3’-5’-DNA exonuclease most likely contributing to the stronger effects in
Act5C> hTREX1 V235G fs
flies.
RVCL-S manifestations increase with aging leading to premature death (
2
). Comparing two
Drosophila
age groups (1 week and 5 weeks after eclosure), we noticed the vessel deterioration in older flies carrying full length hTREX1 compared to the ‘no transgene’ controls (
Fig.3A-B
and
Fig. 4E
). Similar observations were made by Chauvin et al. (2024) (
11
) when hTREX1 was expressed in
D. melanogaster
eye tissue and caused a rough phenotype, though less severe that of RVCL-S isoform, but distinct from the normal state. The authors observed hTREX1 nuclear mis localization, however less pronounced than in hTREX1 V235G fs containing cells. The vascular damage in aged hTREX1 carrying flies might occur as a consequence of hTREX1 mis localization to the nucleus by the mechanisms described in (
14
) due to significant mechanical stress within the vessel tissues.
In summary, we have created an experimental methodology aimed to facilitate the development of a treatment for a rare genetic disorder, RVCL-S. Current work represents an interdisciplinary approach, where humans’ medical problems are addressed using
Drosophila
model organism through the methods of genetics, bioinformatics, biophysics and others. Through optimization of these tools, we have built a model that in the future could be used for testing chemical compounds. Our system also allows us to conduct further research on molecular level to identify the druggable gene targets.
Materials and methods
Drosophila stocks
Fly stocks were maintained on standard cornmeal media at room temperature.
y[1] w[*]; P{Act5C-GAL4-w}E1/CyO
driver was obtained from Bloomington Drosophila Stock Center, stock #25374.
cg3165
CDS deletion and GAL4 knock-in was achieved by CRISPR and performed by WellGenetics (Taiwan) resulting in
w
1118
; cg3165
KO
GAL4 loxP RFP loxP/ CyO
stock. To create transgenic stocks carrying human TREX1 gene, we used a plasmid pcDNA3.1 N-HA-human TREX1 (WT) provided by Jonathan Miner (University of Pennsylvania Perelman School of Medicine) as a source. We performed seamless cloning using NEBuilder® HiFi DNA Assembly Master Mix (NEB, E2621S). HA-hTREX1 DNA fragment was amplified with 5’-GGG AAT TGG GAA TTC GTT AAC ACT AGC GTT TAA ACT TAA GCT TGC CAC CAT GTA CCC-3’ and 5’-ATC CTC TAG AGG TAC CCG CGG CCG CCA CTG TGC T-3’ primers and assembled with pUAST-attB (#1419, DGRC) cut with
Xho
I,
Bgl
II. To create transgenic stocks carrying human TREX1 G235V fs gene we used a plasmid pcDNA3.1 hTREX1 V235Gfs C-HA provided by Jonathan Miner. hTREX1 V235Gfs C-HA DNA fragment was amplified with 5’-TAG GGA ATT GGG AAT TCG TTA ACA CTA GCC ACC ATG GG-3’ and 5’-ATC CTC TAG AGG TAC CCA GCG GGT TTA TCA AGC GTA AT-3’ and assembled with pUAST-attB/
Xho
I,
Bgl
II. Resulting
Drosophila
transformation vectors were microinjected into fly embryos by BestGene Inc. Phi31 mediated cassette exchange occurred at attP2 landing pad. Transgenes integration was verified by PCR and sequencing.
yw; UAS-HA-hTREX1 w
+
y
+t7.7
attP2/TM6B
and
yw; UAS-hTREX1 V325G fs w
+
y
+t7.7
attP2/TM6B
stocks were established.
Genetic crosses
w
1118
; cg3165
KO
GAL4 loxP RFP loxP/ CyO
females were crossed to
yw; UAS-HA-hTREX1 w
+
y
+t7.7
attP2/TM6B
or
yw; UAS-hTREX1 V325G fs w
+
y
+t7.7
attP2/TM6B.
Non-
Cy
, non-
Tb
progeny were collected for the experiments. Non-
Cy
progeny from
w
1118
; cg3165
KO
GAL4 loxP RFP loxP/ CyO
and
yw
cross served as the genetic control.
yw; P{Act5C-GAL4-w}E1/CyO
females were crossed to
yw; UAS-HA-hTREX1 w
+
y
+t7.7
attP2/TM6B Tb.
Non-
Cy
, non-
Tb
progeny was collected for imaging and other experiments. Same crossing scheme was applied for hTREX1 G235V fs carrying flies. Non-
Cy
progeny from
yw; P{Act5C-GAL4-w}E1/CyO
and
yw
cross served as the genetic control.
OCM imaging
Adult flies were imaged using our custom-built SD-OCM system. Broadband light was sent from a super luminescent diode (SLD) with a center wavelength of 850 nm and a bandwidth of 165 nm (Superlum, cBLMD-T-850-HP). A 10x objective focused light onto the sample stage. Light interference from sample and reference arms was measured using a spectrometer and 2048-pixel line-scan camera (Wasatch Photonics, CS800-840/180-80-OC2K-U3). The lateral resolution was measured as ∼2.8 µm and the axial resolution was ∼3.3 µm in tissue. Measured system sensitivity was ∼95.1 dB.
Flies were fixed to a glass slide by attaching their wings to the slide with rubber cement (Elmer’s, Rubber Cement). The OCM beam was positioned over the A1 segment of the heart. M-Mode imaging was performed using 128 A scans and 2000 B scans with an exposure time of 50 µs and a frame rate of 125 frames per second, for an approximately 16-second-long recording per collection.
OCM image processing
Raw OCM data was processed into images using custom lab MATLAB code. To quantify relevant heart parameters, the heart area was segmented in each image using FlyNet3.0 to create masks of the heart area. Once masks were created, they were resized, such that each pixel was equivalent to 1 micron. Heart area over time was plotted, and peak (maximum area) and valley (minimum area) points were identified. Heart rate was calculated using the inverse of the distance between valley areas. End diastolic area (EDA) was calculated as the average area in µm
3
at each peak, and end systolic area (ESA) was calculated as the average area in µm
3
at each valley. Fractional shortening (FS) was calculated using the following equation:
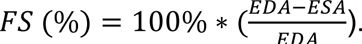
PCR
hTREX1 transgenes were verified by PCR using GoTaq Master Mix (Promega, M7122) and primers 5’-CCT GCA GGTCGGAGT ACT GT-3’ and 5’-GGA AAG TCC TTG GGG TCT TC-3’ following manufacturer’s instructions.
RT-PCR
Total RNA was extracted from adult flies using TRI Reagent (Sigma-Aldrich, T9424-25ML) according to manufacturer’s instructions. cDNA synthesis was performed using QuantiTect Reverse Transcription Kit (Qiagen, 205311) according to the manual. Gene specific PCRs were done using GoTaq Master Mix and primers 5’-AGC GAG ATC ACA GGT CTG AG-3’ and 5’-ACC ACT GCT CCC AT CAT CA-3’ to detect hTREX1 transgenes, and 5’-TGC CCA TTT ATG AGG GCT AC-5’ and 5’-ATC TCC TGC TCG AAG TCC AA-3’ specific for Actin 42A gene serving as a control. qRT-PCR was performed on StepOnePlus System (Applied Biosystems) using QuantiTect SYBR Green PCR Kit (Qiagen, 204143) and hTREX1 specific primers, 5’-
GCATGGGCGTCAATGTTTTG
-3’ and 5’-
TGCTATCCACACAGAAGGCA
-3’. Actin 42A gene served as reference.
Western blots
Total protein extracts were obtained from adult flies as follows: flies were homogenized in 1X Laemmli Sample Buffer (Bio-Rad, 1610737) and boiled for 5 min. Proteins were resolved by size in 10% SDS-PAGE and transferred to nitrocellulose membrane (Bio-Rad, 1620112). Membranes we blocked in 5% Blotto (Santa Cruz Biotechnology, sc-2324) in TBS-T. Anti-HA rMs-IgG1-s (DSHB) antibodies were used at 1:1,000 dilution; anti-LaminC (LC28.26-s, DSHB) at 1:1,000; anti-TREX1 (D8E2O) Rabbit mAb #15107 (Cell Signaling Technology) in 2.5% Blotto. Secondary Peroxidase- conjugated AffiniPure Goat Anti-mouse IgG (Jackson ImmunoResearch, 115-035-003) or Peroxidase-conjugated AffiniPure Goat Anti-rabbit IgG (Jackson ImmunoResearch, 111-035-003) were diluted to 1:100,000. SuperSignal™ West Femto Maximum Sensitivity Substrate (Thermo Scientific, 34095) was used for signal development. Signal visualization was performed on iBright750 imaging system.
Climbing assay
20 flies were placed in an empty vial, left to recover from CO
2
anesthesia for ∼15 min, gently banged down to bring the flies to the bottom of the vial and then flies were let to climb up the wall. Short videos were recorded and used to calculate the percentage of flies crossed the horizontal line drawn at 2.5 cm height at 10 sec. Multiple vials were taped together. Males and females were tested separately. Bar graphs were created in
https://BioRender.com
.
Longevity assay
100 males and 100 females eclosed within 24h were placed in fresh vials, 33-34 flies per vial and kept at 25C 70% humidity. Flies transfer was done every 3-4 days; numbers of dead flies were recorded.
Statistical analyses
For climbing assay and heart function analyses, two-sample student’s t tests were performed with a 95% confidence level. For longevity assay, the age of flies was tracked for each dataset, and we calculated the Kaplan-Meier survival curve, and performed a log-rank test and two-sample two-tailed
t
-test, also with a 95% confidence level.
Data availability
Data underlying the results presented in this paper are not publicly available at this time but may be obtained from the authors upon reasonable request.
Additional information
Funding
This research was supported by National Institutes of Health grant R01-HL156265 (C. Z.); National Institutes of Health grant R01AI143982 (J.J.M.); National Institutes of Health grant R01NS131480 (J.J.M.); Clayco Foundation Innovative Research Award (C. Z.); gift from the Clayco Foundation (J.J.M.); National Science Foundation Graduate Research Fellowship Program (A. Matt); Washington University in St. Louis startup fund (C. Z.); Penn Colton Center for Autoimmunity pilot award (J.J.M.); Penn RVCL Sisters Fund (J.J.M.).
We thank Joanna Chen and Dante Zou for their help with the experiments. We are very grateful to the organizers and participants of the first International RVCL-S Meeting (Leiden, Netherlands, 2024) for helpful and enriching discussions.
Funding
HHS | National Institutes of Health (NIH) (R01-HL156265)
Chao Zhou
HHS | National Institutes of Health (NIH) (R01AI143982)
Jonathan J Miner
HHS | National Institutes of Health (NIH) (R01NS131480)
Jonathan J Miner
Clayco Foundation (Innovative Research Award)
Chao Zhou
Clayco Foundation
Jonathan J Miner
NSF | National Science Foundation Graduate Research Fellowship Program (GRFP)
Abigail Matt
Washington University in St. Louis (WashU) (Startup fund)
Chao Zhou
Penn Colton Center for Autoimmunity pilot award
Jonathan J Miner
Penn RVCL Sisters Fund
Jonathan J Miner
Additional files
Supplementary video 1.
Dorsal vessel crossection of WT (
yw
) male. Yellow scale bar is 50 um.
Supplementary video 2.
Dorsal vessel crossection of
cg3165
KO male. Yellow scale bar is 50 um.
Supplementary video 3.
Dorsal vessel crossection of
cg3165> hTREX1
male. Yellow scale bar is 50 um.
Supplementary video 5.
Dorsal vessel crossection of
Act5C> yw
male. Yellow scale bar is 50 um.
Supplementary video 6.
Dorsal vessel crossection of
Act5C> hTREX1
male. Yellow scale bar is 50 um.
References
1.
Retinal Vasculopathy with Cerebral Leukoencephalopathy and Systemic manifestations (RVCL-S): An update on basic science and clinical perspectives
Cereb Circ Cogn Behav
3
:100046
Google Scholar
2.
Retinal vasculopathy with cerebral leukoencephalopathy and systemic manifestations
Brain
139
:2909–2922
Google Scholar
3.
Retinal Vasculopathy with Cerebral Leukoencephalopathy and Systemic Manifestations
In:
Adam M. P.
et al
, editors.
GeneReviews
Seattle
:
University of Washington
Google Scholar
4.
Cerebroretinal Vasculopathy
Ophthalmology
95
:649–659
Google Scholar
5.
Clinical and genetic analysis of a large Dutch family with autosomal dominant vascular retinopathy, migraine and Raynaud’s phenomenon
Brain
121
:303–316
Google Scholar
6.
Hereditary endotheliopathy with retinopathy, nephropathy, and stroke(HERNS)
Neurology
49
:1322–1330
Google Scholar
7.
Hereditary Vascular Retinopathy, Cerebroretinal Vasculopathy, and Hereditary Endotheliopathy with Retinopathy, Nephropathy, and Stroke Map to a Single Locus on Chromosome 3p21.1-p21.3
The American Journal of Human Genetics
69
:447–453
Google Scholar
8.
C-terminal truncations in human 3′-5′ DNA exonuclease TREX1 cause autosomal dominant retinal vasculopathy with cerebral leukodystrophy
Nat Genet
39
:1068–1070
Google Scholar
9.
NFL and GFAP in (pre)symptomatic RVCL-S carriers: a monogenic cerebral small vessel disease
J Neurol
271
:4138–4145
Google Scholar
10.
Detection of interferon alpha protein reveals differential levels and cellular sources in disease
Journal of Experimental Medicine
214
:1547–1555
Google Scholar
11.
Inherited C-terminal TREX1 variants disrupt homology-directed repair to cause senescence and DNA damage phenotypes in Drosophila, mice, and humans
Nat Commun
15
:4696
Google Scholar
12.
Crizanlizumab for retinal vasculopathy with cerebral leukoencephalopathy in a phase II clinical study
J Clin Invest
134
Google Scholar
13.
Increased Mortality and Vascular Phenotype in a Knock-In Mouse Model of Retinal Vasculopathy With Cerebral Leukoencephalopathy and Systemic Manifestations
Stroke
51
:300–307
Google Scholar
14.
Compromised nuclear envelope integrity drives TREX1-dependent DNA damage and tumor cell invasion
Cell
184
:5230–5246
Google Scholar
15.
The power of Drosophila in modeling human disease mechanisms
Disease Models & Mechanisms
15
:dmm049549
Google Scholar
16.
Drosophila as a model for the identification of genes causing adult human heart disease
Proc. Natl. Acad. Sci. U.S.A.
103
:1394–1399
Google Scholar
17.
Drosophila tools and assays for the study of human diseases
Disease Models & Mechanisms
9
:235–244
Google Scholar
18.
Updates to the Alliance of Genome Resources central infrastructure
Genetics
227
:iyae049
Google Scholar
19.
Diversity and dynamics of the Drosophila transcriptome
Nature
512
:393–399
Google Scholar
20.
Manual annotation of Drosophila genes: a Genomics Education Partnership protocol
F1000Res
11
:1579
Google Scholar
21.
CrossMap: a versatile tool for coordinate conversion between genome assemblies
Bioinformatics
30
:1006–1007
Google Scholar
22.
Measuring TREX1 and TREX2 exonuclease activities
Methods Enzymol
625
:109–133
Google Scholar
23.
Cisplatin depletes TREX2 and causes Robertsonian translocations as seen in TREX2 knockout cells
Cancer Res
67
:9077–9083
Google Scholar
24.
Suppression of TREX1 deficiency-induced cellular senescence and interferonopathies by inhibition of DNA damage response
iScience
26
:107090
Google Scholar
25.
DeepTMHMM predicts alpha and beta transmembrane proteins using deep neural networks
http://biorxiv.org/lookup/doi/10.1101/2022.04.08.487609
26.
Drosophila, an emerging model for cardiac disease
Gene
342
:1–11
Google Scholar
27.
Threat induces cardiac and metabolic changes that negatively impact survival in flies
Current Biology
31
:5462–5472
Google Scholar
28.
Optogenetic pacing in Drosophila melanogaster
Sci. Adv
1
:e1500639
Google Scholar
29.
Dual color optogenetic tool enables heart arrest, bradycardic, and tachycardic pacing in Drosophila melanogaster
Commun Biol
7
:1056
Google Scholar
30.
Developing Drosophila melanogaster Models for Imaging and Optogenetic Control of Cardiac Function
JoVE
63939
https://doi.org/10.3791/63939
Google Scholar
31.
Non-invasive red-light optogenetic control of Drosophila cardiac function
Commun Biol
3
:336
Google Scholar
32.
Drosophila Preparation and Longitudinal Imaging of Heart Function In Vivo Using Optical Coherence Microscopy (OCM)
Journal of Visualized Experiments : JoVE
55002
https://doi.org/10.3791/55002
Google Scholar
33.
FlyNet 2.0: drosophila heart 3D (2D + time) segmentation in optical coherence microscopy images using a convolutional long short-term memory neural network
Biomed Opt Express
11
:1568
Google Scholar
34.
Segmentation of Drosophila heart in optical coherence microscopy images using convolutional neural networks
Journal of Biophotonics
11
:e201800146
Google Scholar
35.
A Drosophila heart optical coherence microscopy dataset for automatic video segmentation
Sci Data
10
:886
Google Scholar
36.
An Attention LSTM U-Net model for Drosophila melanogaster heart tube segmentation in optical coherence microscopy images
Biomed Opt Express
15
:3639–3653
Google Scholar
37.
Targeted gene expression as a means of altering cell fates and generating dominant phenotypes
Development
118
:401–415
Google Scholar
38.
A de novo p.Asp18Asn mutation in TREX1 in a patient with Aicardi–Goutières syndrome
American J of Med Genetics Pt A
152A
:2612–2617
Google Scholar
39.
High-fidelity promoter profiling reveals widespread alternative promoter usage and transposon-driven developmental gene expression
Genome Res
23
:169–180
Google Scholar
40.
Conserved noncoding transcription and core promoter regulatory code in early Drosophila development
eLife
6
:e29005
https://doi.org/10.7554/eLife.29005
Google Scholar
41.
Construction of Transgenic Drosophila by Using the Site-Specific Integrase From Phage C31
Genetics
166
:1775–1782
Google Scholar
42.
Drosophila as a model to study cardiac aging
Experimental gerontology
46
:326
Google Scholar
43.
Age-Associated Cardiac Dysfunction in Drosophila melanogaster
Circulation Research
88
:1053–1058
Google Scholar
44.
Flight and Climbing Assay for Assessing Motor Functions in Drosophila
Bio Protoc
8
:e2742
Google Scholar
45.
Mitochondrial Neurodegeneration: Lessons from Drosophila melanogaster Models
Biomolecules
13
:378
Google Scholar
46.
Misdirected yet intact TREX1 exonuclease activity causes human cerebral and systemic small vessel disease
Brain
https://doi.org/10.1093/brain/awaf085
Google Scholar
Article and author information
Author information
Cite all versions
You can cite all versions using the DOI
https://doi.org/
10.7554/eLife.109595
. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2026,
Gracheva et al.
This article is distributed under the terms of the
Creative Commons Attribution License
, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
views
0
downloads
0
citations
0
Views, downloads and citations are aggregated across all versions of this paper published by eLife.

